Math Is Fun Forum
You are not logged in.
- Topics: Active | Unanswered
#2576 2025-08-27 19:19:38
- Jai Ganesh
- Administrator

- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2376) Shinkansen
Gist
The Shinkansen, or "Bullet Train," is Japan's high-speed railway network, known for its speed, reliability, and advanced technology. Opened in 1964 to connect distant regions and facilitate economic growth, the Shinkansen network allows passengers to travel efficiently between major cities like Tokyo, Osaka, and Fukuoka. The trains are characterized by their aerodynamic, bullet-shaped noses and operate on dedicated tracks, with a central management system ensuring high levels of safety.
The Shinkansen, or "Bullet Train," is Japan's high-speed railway network, known for its speed, reliability, and advanced technology. Opened in 1964 to connect distant regions and facilitate economic growth, the Shinkansen network allows passengers to travel efficiently between major cities like Tokyo, Osaka, and Fukuoka. The trains are characterized by their aerodynamic, bullet-shaped noses and operate on dedicated tracks, with a central management system ensuring high levels of safety.
Summary
The Shinkansen, colloquially known in English as the bullet train, is a network of high-speed railway lines in Japan. It was initially built to connect distant Japanese regions with Tokyo, the capital, to aid economic growth and development. Beyond long-distance travel, some sections around the largest metropolitan areas are used as a commuter rail network. It is owned by the Japan Railway Construction, Transport and Technology Agency and operated by five Japan Railways Group companies.
Starting with the Tokaido Shinkansen (515.4 km; 320.3 mi) in 1964, the network has expanded to consist of 2,951.3 km (1,833.9 mi) of lines with maximum speeds of 260–320 km/h (160–200 mph), 283.5 km (176.2 mi) of Mini-shinkansen lines with a maximum speed of 130 km/h (80 mph), and 10.3 km (6.4 mi) of spur lines with Shinkansen services. The network links most major cities on the islands of Honshu and Kyushu, and connects to Hakodate on the northern island of Hokkaido. An extension to Sapporo is under construction and was initially scheduled to open by fiscal year 2030, but in December 2024, it was delayed until the end of FY2038. The maximum operating speed is 320 km/h (200 mph) (on a 387.5 km (241 mi) section of the Tōhoku Shinkansen). Test runs have reached 443 km/h (275 mph) for conventional rail in 1996, and up to a world record 603 km/h (375 mph) for SCMaglev trains in April 2015.
The original Tokaido Shinkansen, connecting Tokyo, Nagoya, and Osaka —three of Japan's largest cities — is one of the world's busiest high-speed rail lines. In the one-year period preceding March 2017, it carried 159 million passengers, and since its opening more than six decades ago, it has transported more than 6.4 billion total passengers. At peak times, the line carries up to 16 trains per hour in each direction with 16 cars each (1,323-seat capacity and occasionally additional standing passengers) with a minimum headway of three minutes between trains.
The Shinkansen network of Japan had the highest annual passenger ridership (a maximum of 353 million in 2007) of any high-speed rail network until 2011, when the Chinese high-speed railway network surpassed it at 370 million passengers annually.
Details
Shinkansen, pioneer high-speed passenger rail system of Japan, with lines on the islands of Honshu, Kyushu, and Hokkaido. It was originally built and operated by the government-owned Japanese National Railways and has been part of the private Japan Railways Group since 1987.
The first section of the original line, a 320-mile (515-km) stretch between Tokyo and Ōsaka, was opened in 1964. Known as the New Tōkaidō Line, it generally follows and is named for the historic and celebrated Tōkaidō (“Eastern Sea Road”) highway that was used especially during the Edo (Tokugawa) period (1603–1867). Inauguration of the line, just before the start of the Tokyo 1964 Olympic Games, was greeted by widespread international acclaim, and the Shinkansen was quickly dubbed the “bullet train” for the great speed the trains obtained and for the aerodynamic bullet shape of their noses. Many innovations, such as the use of prestressed concrete ties and mile-long welded sections of track, were introduced in the line’s construction. A 100-mile (160-km) extension of that line westward from Ōsaka to Okayama was completed in 1972, and its final segment, a 244-mile (393-km) stretch between Okayama and the Hakata station in Fukuoka, northern Kyushu, opened in 1975.
Other lines radiating northward from Tokyo were completed in 1982 to the cities of Niigata (the Jōetsu line) and Morioka (the Tōhoku line), the Tōhoku line subsequently being extended northward to Hachinohe in 2002. Work to build a link to Aomori, northwest of Hachinohe, began in the late 1990s. When that segment opened in 2010, the Shinkansen was essentially complete for the entire length of Honshu. However, plans had long been in place to connect all three main Japanese islands by Shinkansen with a line northward into Hokkaido (via the Seikan Tunnel under Tsugaru Strait). Construction on the Hokkaido line began in 2005 on the segment between Aomori and Hakodate in southern Hokkaido, the ultimate goal being to extend the line to Sapporo. The line between Aomori and Hakodate opened in 2016. Construction on the section from Hakodate to Sapporo was begun in 2012 and expected to be completed in 2031.
Branches from the Tōhoku line to Yamagata opened in 1992 (extended north to Shinjo in 1999) and to Akita in 1997; a branch from the Jōetsu line to Nagano also opened in 1997. Segments of a further extension of the Nagano branch westward to Toyama and Kanazawa opened in 2015. In addition, a line was completed between Yatsushiro and Kagoshima in southwestern Kyushu in 2004. In the late 1990s work commenced to extend that line northward from Yatsushiro to Hakata, and the opening of the segment in 2011 completed the full north-south route of the Shinkansen on Kyushu. Work began in 2008 on a branch from the Kyushu line southwestward to Nagasaki, and it opened in 2022.
Much of the system’s track runs through tunnels, including one under Shimonoseki Strait between Honshu and Kyushu, another on the Tokyo-Niigata line that is 14 miles (23 km) long, and another near Aomori with a record length (for a double-tracked inland tunnel) of 16.5 miles (26.5 km) when the bore was finished in 2005. Several hundred trains operate daily on the Shinkansen system. The most-frequent service is between Tokyo and Ōsaka, especially during the morning and evening rush hours, when trains depart at intervals of 10 minutes or less. The fastest trains can make the trip from Tokyo to Hakata in about five hours, and the quickest from Tokyo to Aomori take about three hours.
The electric multiple-unit trains, which can seat 1,000 passengers or more, derive their power from an overhead wire system. Trains originally reached top speeds of 130 miles (210 km) per hour, but improvements in track, train cars, and other components have made possible maximum speeds of between 150 and 185 miles (240 and 300 km) per hour. In early 2013 some trains began operating at up to 200 miles (320 km) per hour. Such high speeds made it necessary to develop elaborate safety features. Each car, for example, is equipped with brakes consisting of cast-iron discs and metallic pad linings specially designed not to distort under emergency braking. Moreover, all movements of the trains are monitored and controlled by a central computerized facility in Tokyo.
Additional Information
Japan's main islands of Honshu, Kyushu and Hokkaido are served by a network of high speed train lines that connect Tokyo with most of the country's major cities. Japan's high speed trains (bullet trains) are called shinkansen and are operated by Japan Railways (JR).
Running at speeds of up to 320 km/h, the shinkansen is known for punctuality (most trains depart on time to the second), comfort (relatively silent cars with spacious, always forward-facing seats), safety (no fatal accidents in its history) and efficiency. Thanks to various rail passes, the shinkansen can also be a cost-effective means of travel.
Shinkansen network
The shinkansen network consists of multiple lines, among which the Tokaido Shinkansen (Tokyo - Nagoya - Kyoto - Osaka) is the oldest and most popular. All shinkansen lines (except the Akita and Yamagata Shinkansen) run on tracks that are exclusively built for and used by shinkansen trains. Most lines are served by multiple train categories, ranging from the fastest category that stops only at major stations to the slowest category that stops at every station.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2577 2025-08-28 17:59:13
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2377) Chimpanzee
Gist
Chimpanzees are highly intelligent primates, demonstrating problem-solving skills, tool use, complex communication, and social intelligence. They can learn to use symbols, understand basic commands, and even display empathy and cultural behaviors. While their brains are smaller than humans, they exhibit remarkable cognitive abilities within their own context.
While chimpanzees can form affectionate bonds and exhibit friendly behavior, their unpredictability means that they can also be dangerous. Their wild instincts, intelligence, and emotional depth make them fascinating, but they also deserve respect. Chimpanzee are not pets and should not be treated as such.
Summary
The chimpanzee, also simply known as the chimp, is a species of great ape native to the forests and savannahs of tropical Africa. It has four confirmed subspecies and a fifth proposed one. When its close relative the bonobo was more commonly known as the pygmy chimpanzee, this species was often called the common chimpanzee or the robust chimpanzee. The chimpanzee and the bonobo are the only species in the genus Pan. Evidence from fossils and DNA sequencing shows that Pan is a sister taxon to the human lineage and is thus humans' closest living relative.
The chimpanzee is covered in coarse black hair but has a bare face, fingers, toes, palms of the hands, and soles of the feet. It is larger and more robust than the bonobo, weighing 40–70 kg (88–154 lb) for males and 27–50 kg (60–110 lb) for females and standing 150 cm (4 ft 11 in).
The chimpanzee lives in groups that range in size from 15 to 150 members, although individuals travel and forage in much smaller groups during the day. The species lives in a strict male-dominated hierarchy, where disputes are generally settled without the need for violence. Nearly all chimpanzee populations have been recorded using tools, modifying sticks, rocks, grass and leaves and using them for hunting and acquiring honey, termites, ants, nuts and water. The species has also been found creating sharpened sticks to spear small mammals. Its gestation period is eight months. The infant is weaned at about three years old but usually maintains a close relationship with its mother for several years more.
The chimpanzee is listed on the IUCN Red List as an endangered species. Between 170,000 and 300,000 individuals are estimated across its range. The biggest threats to the chimpanzee are habitat loss, poaching, and disease. Chimpanzees appear in Western popular culture as stereotyped clown-figures and have featured in entertainments such as chimpanzees' tea parties, circus acts and stage shows. Although chimpanzees have been kept as pets, their strength, aggressiveness, and unpredictability makes them dangerous in this role. Some hundreds have been kept in laboratories for research, especially in the United States. Many attempts have been made to teach languages such as American Sign Language to chimpanzees, with limited success.
Details
A chimpanzee, (Pan troglodytes), is a species of ape that, along with the bonobo, is most closely related to humans. Chimpanzees inhabit tropical forests and savannas of equatorial Africa from Senegal in the west to Lake Albert and northwestern Tanzania in the east. Individuals vary considerably in size and appearance, but chimpanzees stand approximately 1–1.7 metres (3–5.5 feet) tall when erect and weigh about 32–60 kg (70–130 pounds). Males tend to be larger and more robust than females. Chimpanzees are covered by a coat of brown or black hair, but their faces are bare except for a short white beard. Skin colour is generally white except for the face, hands, and feet, which are black. The faces of younger animals may be pinkish or whitish. Among older males and females, the forehead often becomes bald and the back becomes gray.
Natural history
Chimpanzees awaken at dawn, and their day is spent both in the trees and on the ground. After a lengthy midday rest, late afternoon is usually the most intensive feeding period. In the trees, where most feeding takes place, chimps use their hands and feet to move about. They also leap and swing by their arms (brachiate) skillfully from branch to branch. Movement over any significant distance usually takes place on the ground. Though able to walk upright, chimpanzees more often move about on all fours, leaning forward on the knuckles of their hands (knuckle walking). At night they usually sleep in the trees in nests they build of branches and leaves. Chimpanzees are unable to swim, but they will wade in water. The chimpanzee diet is primarily vegetarian and consists of more than 300 different items, mostly fruits, berries, leaves, blossoms, and seeds but also bird eggs and chicks, many insects, and occasionally carrion. Chimpanzees also hunt, both alone and in groups, stalking and killing various mammals such as monkeys, duikers, bushbucks, and wild pigs. They also appear to use certain plants medicinally to cure diseases and expel intestinal parasites.
The female chimpanzee bears a single young at any time of year after a gestation period of about eight months. The newborn weighs about 1.8 kg (about 4 pounds), is almost helpless, and clings to the fur of the mother’s belly as she moves. From about 6 months to 2 years, the youngster rides on the mother’s back. Weaning takes place at about 5 years. Males are considered adults at 16 years of age, and females usually begin to reproduce at about 13 years, but often only two offspring survive during her lifetime. The longevity of chimps is about 45 years in the wild and 58 in captivity; however, older individuals have been documented. For example, Cheetah the chimpanzee, an animal actor from the Tarzan movies of the 1930s and ’40s, was reported to have lived approximately 80 years.
Conservation status
Chimpanzees are an endangered species; their population in the wild has been reduced by hunting (primarily for meat), destruction of habitat from logging or farming, and commercial exportation for use in zoos and research laboratories. The International Union for Conservation of Nature (IUCN) noted that, despite having one of the largest geographic ranges of the great apes, chimpanzee populations have fallen significantly since the 1980s. Lions and leopards also prey upon chimpanzees.
Social behaviour
Chimpanzees are lively animals with more extraverted dispositions than either gorillas or orangutans. They are highly social and live in loose and flexible groups known as communities, or unit groups, that are based on associations between adult males within a home range, or territory. Home ranges of forest-dwelling communities can be as small as a few square kilometres, but home ranges covering hundreds of square kilometres are known among savanna communities. A community can number from 20 or fewer to well over 100 members. Each consists of several subgroups of varying size and unstable composition. Social dominance exists, with adult males being dominant over adult females and adolescent males. Within a community, there are twice or three times as many adult females as adult males; the number of adults is about equal to the number of immature individuals. Communities usually divide into subgroups called parties, which vary widely in size. The dominance hierarchy among male chimpanzees is very fluid; individuals associate with each other and join and leave different subgroups with complete freedom. The dominant (alpha) male of a group can monopolize ovulating females through possessive behaviour. On the other hand, gang attack by subordinate males can expel an alpha male. Males spend all of their lives in the community they are born in, but occasionally a juvenile male may transfer to another community with his mother. In contrast to males, most females leave their group of birth to join a neighbouring group when they mature at around age 11. Female chimpanzees spend most of their time with their young or with other females. Those with dependent offspring are more likely to range alone or in small parties within narrow “core areas.” Females have been known to form coalitions against a bullying adult male or newly immigrated female.
Relations between different chimp communities tend to be hostile. Intruders on a group’s home range may be attacked, and adult males engage in boundary patrol. On rare occasions, a group may invade a neighbouring territory that is much smaller in size, and fatalities among the smaller group result. Infanticide and cannibalism by adult males, and to a lesser extent by adult females, have been observed. Victimized infants are not only those of neighbouring groups but also those born to newly immigrated females. Between- and within-group competition among individuals of the same sex is the likely cause of such violence. Sometimes a male and female will form a consortship, engaging in exclusive mating relationships by leaving other members of the group and staying in the periphery of the group range. This strategy, however, brings increased risk of attack by neighbouring groups.
Chimpanzees exhibit complex social strategies such as cooperation in combat and the cultivation of coalitions and alliances via ranging together, reciprocal grooming, and the sharing of meat (sometimes in exchange for mating opportunities). An alpha male, for instance, may interfere with his rival in grooming with a third party because such a coalition might jeopardize the alpha’s status. On the other hand, the third party might show strategic opportunism in such a situation, since his assistance to either side could determine which of his superiors prevails. Chimpanzees, therefore, appear to have some concept of “trade.” They console, reconcile, and retaliate during fighting and so share emotions and aspects of psychology similar to those found in humans: self-recognition, curiosity, sympathy, grief, and attribution. Although chimps take care of orphaned infants, they also tease handicapped individuals, conceal information that would bring disadvantage to themselves, and manipulate others for their own advantage by expressing deceptive postures, gestures, and facial expressions.
Intelligence
Chimpanzees are highly intelligent and are able to solve many kinds of problems posed to them by human trainers and experimenters. A number of researchers have taught chimpanzees to use sign language or languages based on the display of tokens or pictorial symbols. The implications of these language studies have been contested, however. Critics charge that apes have not acquired true language in the sense of understanding “words” as abstract symbols that can be combined in meaningful new ways. Other investigators maintain that more recent language training has resulted in the chimpanzees’ acquiring a true recognition of “words” as abstractions that can be applied in novel contexts.
Communication between chimps in the wild takes the form of facial expressions, gestures, and a large array of vocalizations, including screams, hoots, grunts, and roars. Males display excitement by standing erect, stamping or swaying, and letting out a chorus of screams. Chimps use louder calls and gestures for long-distance communication (such as drumming on tree buttresses) and quieter calls and facial expressions for short-distance communication. Similarities to human laughter and smiling might be seen in their “play panting” and grinning, respectively.
Various tools are used in several contexts. Chimpanzees “fish” for termites and ants with probes made of grass stalks, vines, branches, peeled bark, and midribs of leaves. They crack hard nuts open by using stones, roots, and wood as hammers or anvils, and they use “leafy sponges” (a handful of folded leaves or moss) to drink water. Branches and leaves are detached and displayed during courtship. In threat displays, chimps throw rocks and drag and throw branches. Sticks are used to inspect dead pythons or other unfamiliar objects that might be dangerous. Leaves are used hygienically in wiping the mouth or other soiled body parts. Chimpanzees also use different tools in succession as a “tool set.” For example, chimpanzees of the Congo basin first dig into termite mounds with a stout stick and then fish for individual termites with a long, slender wand. Tools are also used in combination as “tool composites.” Chimpanzees in the Guinea region push leafy sponges into hollows of trees containing water and then withdraw the wet sponges by using sticks. Chimps thus differ locally in their repertoire of tool use, with younger animals acquiring tool-using behaviours from their elders. Such cultural differences are also seen in food items consumed and in gestural communication. Chimpanzees indeed possess culture when it is defined as the transmission of information from generation to generation via social learning shared by most members of a single age or sex class in a given group.
Chimpanzees’ intelligence, responsiveness, and exuberance have made them ideal nonhuman subjects for psychological, medical, and biological experiments. Young chimpanzees can become very attached to their human trainers, and their expressions of feeling resemble those of humans more closely than any other animal.
Taxonomy
Genetic analysis suggests that the lineages leading to modern humans and chimpanzees diverged from each other between 6.5 million and 9.3 million years ago and that at least 98 percent of the human and chimpanzee genomes are identical. Chimpanzees are classified taxonomically as a single species, Pan troglodytes. (The so-called pygmy chimpanzee, or bonobo, is a distinct and separate species, P. paniscus, that diverged from chimpanzees about 1.7 million years ago.) Four subspecies of P. troglodytes are recognized: the tschego, or Central African chimpanzee (P. troglodytes troglodytes), also known as the common chimpanzee in continental Europe; the West African, or masked, chimpanzee (P. troglodytes verus), known as the common chimpanzee in Great Britain; the East African, or long-haired, chimpanzee (P. troglodytes schweinfurthii); and the Nigerian-Cameroon chimpanzee (P. troglodytes ellioti, which was formerly classified as P. troglodytes vellerosus).
Additional Information
Like us, chimps are highly social animals, care for their offspring for years and can live to be over 50. In fact, chimpanzees are our closest cousins; we share about 98% of our genes.
In their habitat in the forests of Central Africa, chimpanzees spend most of their days in the treetops. When they do come down to earth, chimps usually travel on all fours, though they can walk on their legs like humans for as far as a mile. They use sticks to fish termites out of mounds and bunches of leaves to sop up drinking water.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2578 2025-09-02 20:18:55
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2378) Rafflesia arnoldii
Gist
Often called the corpse flower, Rafflesia arnoldi blooms into the single largest individual flower in the world. When it does, it emits a vile aroma, similar to rotten meat, attracting insects, such as flies and beetles, that feed on dead flesh.
Summary
Rafflesia is (genus Rafflesia), genus of about 42 species of parasitic plants native to Southeast Asia. All Rafflesia species parasitize the roots of Tetrastigma vines (family Vitaceae), and their presence is not made known until the striking flowers emerge from the host vine. One species, Rafflesia arnoldii, boasts the largest single flower of any plant.
Taxonomy
Kingdom: Plantae
Clade: Angiosperm
Order: Malpighiales
Family: Rafflesiaceae
Genus: Rafflesia
Physical description
Like other members of the family Rafflesiaceae, Rafflesia plants are endoparasitic, meaning that the vegetative organs are so reduced and modified that the plant body exists only as a network of threadlike cellular strands living almost wholly within the tissues of the host plant. There are no green photosynthetic tissues, leaves, roots, or stems in the generally accepted sense, although vestiges of leaves exist in some species as scales. Rafflesia plants are thus obligate parasites, which cannot live without the nutrition provided by the host. Despite the dramatic reduction of most of the plant body, the flowers are well developed and can be extremely large.
Rafflesia flowers are sturdy, typically with five substantial tepals (undifferentiated petals and sepals) surrounding the sex organs in a central cup. Interestingly, individual flowers are unisexual, meaning that pollination can occur only if another flower of the opposite sex is simultaneously blooming. The distinctive flowers are sapromyiophilous, meaning that they are pollinated by carrion-feeding flies, and they have a number of adaptations to attract their unconventional pollinators. Most are reddish or purplish brown and have a mottled pattern that resembles rotting flesh. In addition, they emit a fetid carrion odor during the few days they are blooming, and some species even generate heat to simulate decomposition. The unusual pollen is transferred by means of a sticky liquid that dries on the flies. The resultant fruit is a berry containing sticky seeds thought to be disseminated by fruit-eating rodents.
Major species and conservation
The genus includes the giant R. arnoldii, sometimes known as the corpse flower or monster flower, which produces the largest-known individual flower of any plant species in the world and is found in the forested mountains of Sumatra and Borneo. Its fully developed flower appears aboveground as a thick fleshy five-lobed structure weighing up to 11 kg (24 pounds) and measuring almost one meter (about one yard) across.
Most Rafflesia species are considered rare and, given their complete dependence on Tetrastigma vines, are extremely difficult to cultivate and very vulnerable to extinction. Major threats include the loss of rainforest habitat, due to logging and other land-use changes, and illegal harvest of the flowers for their purported medicinal properties. As of 2023 only one species, R. magnifica of the Philippines, has been formally evaluated, and it is listed as critically endangered by the IUCN Red List of Threatened Species, though scientists estimate that at least 60 percent of the species are endangered.
Details
Kingdom: Plantae
Clade: Tracheophytes
Clade: Angiosperms
Clade: Eudicots
Clade: Rosids
Order: Malpighiales
Family: Rafflesiaceae
Genus: Rafflesia
Species: R. arnoldii
Binomial name
Rafflesia arnoldii, the corpse flower, or giant padma, is a species of flowering plant in the parasitic genus Rafflesia within the family Rafflesiaceae. It is noted for producing the largest individual flower on Earth. It has a strong and unpleasant odour of decaying flesh. It is native to the rainforests of Sumatra and Borneo. Although there are some plants with larger flowering organs like the titan arum (Amorphophallus titanum) and talipot palm (Corypha umbraculifera), those are technically clusters of many flowers.
Rafflesia arnoldii is one of the three national flowers in Indonesia, the other two being the white jasmine (Jasminum sambac) and moon orchid (Phalaenopsis amabilis). It was officially recognised as a national "rare flower" (Indonesian: puspa langka) in Presidential Decree No. 4 in 1993.
Taxonomy
The first European to find Rafflesia was the ill-fated French explorer Louis Auguste Deschamps. He was a member of a French scientific expedition to Asia and the Pacific, detained by the Dutch for three years on the Indonesian island of Java, where, in 1797, he collected a specimen, which was probably what is now known as R. patma. During the return voyage in 1798, his ship was taken by the British, with whom France was at war, and all his papers and notes were confiscated. Joseph Banks is said to have agitated for the return of the stolen documents, but apparently to no avail; they were lost, turned up for sale around 1860, went to the British Museum of Natural History, where they were promptly lost again. They did not see the light of day until 1954, when they were rediscovered at the Museum. To everyone's surprise, his notes and drawings indicate that he had found and studied the plants long before the British. It is thought quite possible the British purposely hid Deschamps' notes, to claim the 'glory' of 'discovery' for themselves.
In 1818 the British surgeon Joseph Arnold collected a specimen of another Rafflesia species found by a Malay servant in a part of Sumatra, then a British colony called British Bencoolen (now Bengkulu), during an expedition run by the recently appointed Lieutenant-Governor of Bencoolen, Stamford Raffles. Arnold contracted a fever and died soon after the discovery, the preserved material being sent to Banks. Banks passed on the materials, and the honour to study them was given to Robert Brown. The British Museum's resident botanical artist Franz Bauer was commissioned to make illustrations of the new plants. Brown eventually gave a speech before the June 1820 meeting of the Linnean Society of London, where he first introduced the genus and its until then two species. Brown gave the generic name Rafflesia in honour of Raffles. Bauer completed his pictures some time in mid-1821, but the actual article on the subject continued to languish.
William Jack, Arnold's successor in the Sumatran Bencoolen colony, recollected the plant and was the first to officially describe the new species under the name R. titan in 1820. It is thought quite likely that Jack rushed the name to publication because he feared that the French might publish what they knew of the species, and thus rob the British of potential 'glory'. Apparently aware of Jack's work, Brown finally had the article published in the Transactions of the Linnean Society a year later, formally introducing the name R. arnoldii (he ignores Jack's work in his article).
Because Jack's name has priority, R. arnoldii should technically be a synonym of R. titan, but at least in Britain, it was common at the time to recognise the names introduced by well-regarded scientists such as Brown, over what should taxonomically be the correct name. This was pointed out by the Dutch Rafflesia expert Willem Meijer in his monographic addition to the book series Flora Malesiana in 1997. Instead of sinking R. arnoldii into synonymy, however, he declared that the name R. titan was "incompletely known": the plant material used by Jack to describe the plant has been lost.
In 1999, the British botanical historian David Mabberley, in response to Meijer's findings, attempted to rescue Brown's names from synonymy. This is known as 'conservation' in taxonomy, and normally this requires making a formal proposal to the committee of the International Code of Botanical Nomenclature (ICBN). Mabberley thought he found a loophole around such a formal review by noting that while Brown was notoriously slow to get his papers published, he often had a handful of pre-print pages privately printed to exchange with other botanists: one of these pre-prints had been recently bought by the Hortus Botanicus Leiden, and it was dated April 1821. Mabberley thus proposed that this document be considered the official effective publication, stating this would invalidate Jack's earlier name. For some reason Mabberley uses 1821, a few months after Brown's pre-print, as the date of Jack's publication, instead of the 1820 publication date in Singapore. Confusingly, the record in the International Plant Names Index (IPNI) still has yet another date, "1823?", as it was in the Index Kewensis before Meijer's 1997 work. Mabberley's proposals regarding Brown's name were accepted by institutions, such as the Index Kewensis.
Mabberley also pointed out that the genus Rafflesia was thus first validated by an anonymous report on the meeting published in the Annals of Philosophy in September 1820 (the name was technically an unpublished nomen nudum until this publication). Mabberley claimed the author was Samuel Frederick Gray. However, as that is nowhere stated in the Annals, per Article 46.8 of the code of ICBN, Mabberley was wrong to formally ascribe the validation to Gray. The validation of the name was thus attributed to one Thomas Thomson, the editor of the Annals in 1820, by the IPNI. Mabberley admitted his error in 2017. This Thomson was not the botanist Thomas Thomson, who was three years old in 1820, but his identically named father, a chemist, and Rafflesia is thus the only botanical taxon this man ever published.
Errata
An old Kew webpage claimed that Sophia Hull was present when the specimen was collected and finished the colour drawing that Arnold had started of the plant. It also stated that Brown had originally wanted to call the plant genus Arnoldii.
Regional names
It is called kerubut in Sumatra. In the kecamatan ('district') of Pandam Gadang, it is known as cendawan biriang in the Minangkabau language.
Description
Although Rafflesia is a vascular plant, it lacks any observable leaves, stems or even roots, and does not have chlorophyll. It lives as a holoparasite on vines of the genus Tetrastigma, most commonly T. angustifolium. Similar to fungi, individuals grow as a mass of thread-like strands of tissue completely embedded within and in intimate contact with surrounding host cells from which nutrients and water are obtained. It can only be seen outside the host plant when it is ready to reproduce; the only part of Rafflesia that is identifiable as distinctly plant-like are the flowers, though even these are unusual since they attain massive proportions, are reddish-brown with white spots, and stink of rotting flesh. According to Sandved, the flower opens with a hissing sound.
The flower of Rafflesia arnoldii grows to a diameter of around 1 m (3 ft 3 in),[2] and weighs up to 11 kg (24 lb). According to the Mongabay institution, the single largest R. arnoldii to be measured was 1.14 m (3 ft 9 in) in width. These flowers emerge from very large, cabbage-like, maroon or dark brown buds typically about 30 cm (12 in) wide, but the largest (and the largest flower bud ever recorded) found at Mount Sago, Sumatra in May 1956 was 43 cm (17 in) in diameter. Indonesian researchers often refer to the bud as a 'knop' (knob). According to one source, these buds require 21 months to form. Yet the flowers remain open for only four days.
The plant is native to the rainforest regions of Malaysia, Indonesia, the Philippines, and Thailand.
Additional Information
Often called the corpse flower, Rafflesia arnoldi blooms into the single largest individual flower in the world.
When it does, it emits a vile aroma, similar to rotten meat, attracting insects, such as flies and beetles, that feed on dead flesh.
These flesh-loving creatures pollinate the flower, allowing it to spread through the rainforests of Borneo.
Due to the incredibly specific requirements of the plant, almost no botanical gardens have a Rafflesia arnoldi in cultivation, including Kew.
Rafflesia arnoldi has no leaves, stems or roots, and is a parasitic plant that grows on vines in the genus Tetrastigma.
Plant description
Rafflesia arnoldi lives inside Tetrastigma vines as a mass of fleshy strands which absorb water and nutrients from the host. It grows out of the host plant's bark as brown, cabbage-like buds called knops which bloom over several days. The flowers have five lobes, are reddish-brown with white spots, and grow up to 1m across. They appear for a week, releasing a scent of rotting meat.
Cultural
The flower is an iconic symbol of southeast Asian rainforest, and has been depicted on several Indonesian postage stamps.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2579 2025-09-03 16:53:36
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2379) Bluff (Geography)
Gist
In geography, a bluff is a high, steep slope or cliff that typically overlooks a body of water, formed by the erosion of the outer bank of a river's meander or along a coastline. The term can also refer to a clump of trees, particularly willows or poplars, found on the flat Canadian prairies, which is a different meaning from the geological landform.
Although any cliff with a broad, steep face may be called a bluff, the term bluff refers to a high bank or bold headland with a broad, precipitous, sometimes rounded cliff face. Bluffs can overlook either a plain or a body of water, especially on the outside curve of a stream meander.
Summary
In geography and geology, a cliff or rock face is an area of rock which has a general angle defined by the vertical, or nearly vertical. Cliffs are formed by the processes of weathering and erosion, with the effect of gravity. Cliffs are common on coasts, in mountainous areas, escarpments and along rivers. Cliffs are usually composed of rock that is resistant to weathering and erosion. The sedimentary rocks that are most likely to form cliffs include sandstone, limestone, chalk, and dolomite. Igneous rocks such as granite and basalt also often form cliffs.
An escarpment (or scarp) is a type of cliff formed by the movement of a geologic fault, a landslide, or sometimes by rock slides or falling rocks which change the differential erosion of the rock layers.
Most cliffs have some form of scree slope at their base. In arid areas or under high cliffs, they are generally exposed jumbles of fallen rock. In areas of higher moisture, a soil slope may obscure the talus. Many cliffs also feature tributary waterfalls or rock shelters. Sometimes a cliff peters out at the end of a ridge, with mushroom rocks or other types of rock columns remaining. Coastal erosion may lead to the formation of sea cliffs along a receding coastline.
The British Ordnance Survey distinguishes between cliffs (continuous line along the topper edge with projections down the face) and outcrops (continuous lines along lower edge).
Details:
What Is A Bluff In Geography?
A bluff is a ridge of land that extends into the air. The term is commonly used in geography to refer to high ground. Bluffs are often situated at the foot of steep cliffs and mountains.
A bluff is a steep hill or cliff, especially one that appears close to its base. It can also refer to an area where the slope is so steep that it appears to be an unbroken cliff.
A typical bluff is situated at the foot of steep cliffs and mountains.
Why Is A Bluff Called A Bluff?
A bluff is a cliff, small hill, or other elevation on the side of a mountain or hill, that rises steeply from the ground, usually without trees or shrubs.
Bluffs are called bluff because they appear to be higher than they actually are. From the distance, they seem to be steeper than they actually are.
How Is A Bluff Formed?
How a bluff is formed in geography is a question that has no clear answer. It all depends on the environment that the bluff was formed in and other factors like how deep it is and how much erosion takes place.
When a bluff is created naturally, it forms by the force of wind and water along with the effect of gravity on the earth’s surface. This natural process mainly happens in steep places where there are high elevation changes.
As the slope becomes too steep for the water or wind to erode it, it builds up and forms a bluff.
Bluffs are also formed by the slow movement of glaciers, ice sheets, and permafrost. Glaciers push large amounts of rock off of mountainside through grinding and crushing – this produces large boulders that act as rubble dams blocking the flow of streams.
How To Identify A Bluff In Geography?
Bluffs are natural formations that can be found near water and mountain ranges. They take many forms and typically look like a hill reaching the sky or a mound of rock.
There are many ways to identify a bluff. A bluff is usually steep enough to have a significant decline or incline on both sides of the top.
The bluffs are not just steep but have uneven, sharp sawtooth edges that provide cover for any assailant. The bluff is also made up of large stones that have become cemented together over time and are covered with moss and vegetation, which further provides cover from above.
Bluff is not higher than its surroundings but it grants an advantage to those who use it as cover from above due to the uneven, sharp edges.
Bluffs are also characterized by their lack of vegetation and the presence of large rock formations jutting out from them.
In order to identify a bluff, you need to look at the shape and size. The bluffs should be steep, irregularly shaped.
What Is A Bluff On A Lake Or Beach?
A bluff is a shoreline that plunges abruptly into the water, as seen in some lakes and rivers. In order to prevent erosion from the water, the land behind a bluff is often protected by planting shrubs and trees along the steep slope of the bank.
A bluff can be created naturally or artificially through human efforts such as earth-moving projects, filling of river beds, building dams, or installing rock protections.
Cliff vs Bluff [What Is A Bluff In Geography]
Here are some of the differences between a bluff and a cliff:
Cliff
A cliff is a sharp change in elevation with little or no horizontal level between two points of different elevations. A cliff face is usually vertical or nearly so but can also be almost flat at low elevations.
Such features are typically found in mountainous terrain and may be formed either by erosion or by other types of geological activity.
Cliffs can be considered natural features that stabilize a slope, preventing further erosion and landslides from occurring on the mountainside, but they have also been used for building retaining walls and even as places to live.
Bluff
A bluff is a high point of land that appears to be very steep, but it is actually not.
Bluffs are generally found on the side of a mountain or hill, but sometimes they are found on level ground or even on the seashore.
Bluffs are defined by their steepness, meaning the difference in elevation between the highest point on top of the bluff and its base.
Conclusion: What Is A Bluff In Geography
When you are talking about geography, bluff can be a natural feature, a geological formation, or an archaeological site. These things that are difficult to see from one perspective are easy to spot from another perspective.
Bluff is the name given to a low-lying area of land that is surrounded by steep cliffs, steep slopes, or water. It’s the place you hide in when you don’t want to be seen.
Additional Information
A bluff is a type of broad, rounded cliff. Most bluffs border a river, beach, or other coastal area.
Bluffs may form along a river where it meanders, or curves from side to side. River currents on the outside of the curve erode, or wear away, the lower part of a river bank. No longer supported, the upper part of the bank breaks off, leaving the high wall of a bluff. The 150-meter (500-foot) Great River Bluffs in the U.S. state of Minnesota, for example, were carved by the meanderings of the mighty Mississippi River.
Erosion also produces bluffs along the wider floodplain of a river. Over thousands of years, a meandering river gradually shifts from side to side across its floodplain. Where the meanders, or loops, of the river reach valley walls, the water may carve bluffs. In fact, a “bluff line” defines the outer limits of a river’s floodplain, and is often another name for valley wall. A floodplain’s bluff lines may be steep and narrow, or they may be wide and gentle.
Coastal bluffs are formed through a combination of erosion from wind, sea spray, and crashing waves. These bluffs are often more rugged than their inland counterparts, and are more vulnerable to major erosion. Coastal bluffs, especially those in the Puget Sound region of the U.S. state of Washington, are sometimes called feeder bluffs. The constant erosion of feeder bluffs supplies (feeds) sediment to the beaches and seashore downstream below.
Another sort of coastal bluff is the beach ridge. Beach ridges are formed entirely by waves lapping onshore, pushing sand and sediment up and away from the body of water. Beach ridges run parallel to the shoreline and are often associated with sand dunes. The Indiana Dunes, for example, are extensions of beach ridges formed by the waters of Lake Michigan. At this national lakeshore, tiny bluffs give way to larger dunes, and the ecosystem eventually creates the ideal conditions for an oak forest through the process of plant succession.
Like many types of cliffs, bluffs provide important information about how Earth developed. Scotts Bluff, for instance, rises more than 1,400 meters (4,600 feet) above the North Platte River in the U.S. state of Nebraska. The exposed rock of this national monument allows geologists to peer more than 30 million years into North America’s past. Rock formations at Scotts Bluff preserve the history of ancient volcanic activity nearby, as well as the fossilized presence of prehistoric North American rhinos, tapirs, and even camels.
Life on the Bluff
The summits of many bluffs are bare, rocky outcroppings. However, the harsh environment of bluffs are often vital ecosystems. Tiny organisms called lichen often colonize rocky bluffs, providing vital nutrients for many insects and birds.
Hardy grasses and shrubs can also take root on rocky bluffs. The shallow roots of these plants slow erosion of the bluff, even helping to secure valuable topsoil in some places.
Bluffs provide an ideal nesting spot for fishing birds such as cormorants and kingfishers. Cormorants build their nests on the bare ground of bluffs, bringing sticks and seaweed for construction. Cormorant colonies can grow so large that they take over the entire bluff. Kingfishers don’t nest on top the bluff, but dig a burrow directly in it. They prefer burrows on bluffs with little vegetation, as root systems get in the way of digging.
Bluffs are even home to endangered species. The El Segundo blue butterfly (Euphilotes battoides allyni), for example, is found only in a small bluff ecosystem near the giant Los Angeles International Airport in Southern California.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2580 2025-09-04 14:33:58
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2380) Penguin
Gist
A penguin is a flightless, black-and-white seabird found almost exclusively in the Southern Hemisphere, known for its adaptations to marine life, including flippers for swimming, dense feathers for warmth, and a diet of fish and krill. While most live in cold Antarctic or sub-Antarctic regions, the Galápagos penguin lives near the equator. Penguins are excellent swimmers and divers, using their short, paddle-like wings to propel themselves through the water, and their contrasting dark backs and white bellies provide camouflage.
Penguins primarily live in the Southern Hemisphere, inhabiting cold environments like Antarctica, subantarctic islands, and temperate coasts in countries such as Australia, New Zealand, and South Africa. While most species are found in these southern regions, the Galápagos penguin is an exception, living on islands near the equator in the warm waters off the coast of Ecuador. All penguins require proximity to the ocean for food and to raise their young on land.
Summary
Penguins are a group of aquatic flightless birds from the family Spheniscidae of the order Sphenisciformes. They live almost exclusively in the Southern Hemisphere. Only one species, the Galápagos penguin, is equatorial, with a small portion of its population extending slightly north of the equator (within a quarter degree of latitude). Highly adapted for life in the ocean water, penguins have countershaded dark and white plumage and flippers for swimming. Most penguins feed on krill, fish, squid and other forms of sea life which they catch with their bills and swallow whole while swimming. A penguin has a spiny tongue and powerful jaws to grip slippery prey.
They spend about half of their lives on land and the other half in the sea. The largest living species is the emperor penguin (Aptenodytes forsteri): on average, adults are about 1.1 m (3 ft 7 in) tall and weigh 35 kg (77 lb). The smallest penguin species is the little blue penguin (Eudyptula minor), also known as the fairy penguin, which stands around 30–33 cm (12–13 in) tall and weighs 1.2–1.3 kg (2.6–2.9 lb). Today, larger penguins generally inhabit colder regions, and smaller penguins inhabit regions with temperate or tropical climates. Some prehistoric penguin species were enormous: as tall or heavy as an adult human. There was a great diversity of species in subantarctic regions, and at least one giant species in a region around 2,000 km south of the equator 35 mya, during the Late Eocene, a climate decidedly warmer than today.
Details
A penguin is (order Sphenisciformes), any of 18–21 species of flightless marine birds that live only in the Southern Hemisphere. The majority of species live not in Antarctica but rather between latitudes 45° and 60° S, where they breed on islands. A few penguins inhabit temperate regions, and one, the Galapagos penguin (Spheniscus mendiculus), lives at the Equator.
General features:
The world's deepest-diving birds
The world's deepest-diving birdsEmperor penguins (Aptenodytes forsteri) live in colonies along the coastline of Antarctica. They are capable of diving to depths of about 550 metres (1,800 feet) in search of food, which makes them Earth's deepest-diving birds.
The stocky, short-legged appearance of penguins has endeared them to people worldwide. They range from about 35 cm (14 inches) in height and approximately 1 kg (about 2 pounds) in weight in the blue, or fairy, penguin (Eudyptula minor) to 115 cm (45 inches) and 25 to 40 kg (55 to 90 pounds) in the emperor penguin (Aptenodytes forsteri). Most are black on the back and white below, often with lines of black across the upper breast or spots of white on the head. Colour is rare, being limited to red or yellow irises of the eye in some species; red beaks or feet in a few; yellow brow tufts in the three species of Eudyptes; and orange and yellow on the head, neck, and breast in the emperor and king (A. patagonica) penguins.
The total populations of some species, such as the emperor, are estimated in the hundreds of thousands, but most species of smaller penguins certainly run into the millions. Immense island breeding colonies, some teeming with hundreds of thousands of nesting pairs, represent a large potential food resource, but the economic importance of penguins is negligible. Nineteenth-century whalers and seal hunters visited some colonies for meat and eggs, and a penguin oil industry once took large numbers of the birds. By the early 20th century, however, this exploitation was no longer profitable, and most colonies were left alone or actively protected. Some species are now increasing in numbers, apparently as a result of the mid-20th century’s decimation of Antarctic whales, which compete with penguins for the krill (minute crustaceans) on which both feed. Penguin populations, however, are highly vulnerable to changes in climate and ocean temperature, including recent global warming. Penguins also are very sensitive to depletion of local fish populations by humans.
Natural history:
Reproduction
Many features of the penguin life cycle vary with body size and geographic distribution; the chronology of breeding may also vary within a species in relation to latitude. The majority of species breed only once each year. Certain species, such as the African penguin (Spheniscus demersus), probably other members of this genus, and the blue penguin, breed twice a year. The king penguin breeds twice in three years. One egg is laid by the emperor and king penguins; all others lay two or occasionally three. Most penguins begin breeding in the austral (southern) spring or summer. King penguins are on a 14- to 18-month cycle, and the timing of an individual pair depends on the success or failure of the previous breeding attempt. Some populations of the gentoo penguin (Pygoscelis papua) also breed in winter. The breeding of the emperor penguin begins in autumn, apparently timed so that the long developmental period will produce the young in midsummer, when their chances of survival are greatest.
The gentoo, which has a circumpolar distribution, is notable for its lack of synchrony among populations, but otherwise its breeding schedule is essentially comparable to that of most other species. In the Crozet Islands off southern Africa, for example, egg laying takes place in July. The two eggs are incubated for 35 or 36 days, and the rearing of the chick takes two months. The last immature birds go to sea in January.
Many types of visual and vocal displays are employed between the arrival of the birds at the colony and their departure. Courtship calls are used during pairing and to a lesser degree during the succeeding phases of breeding. There are marked vocal differences between sexes in the emperor penguin and the king penguin and less-marked dimorphisms in some other species. Upon arrival at the colony each bird returns to the nest that it left the previous year and generally rejoins its mate of the previous year, unless the death of the latter forces it to choose another partner. This applies even to the emperor penguin, which is capable of finding its mate despite the absence of a nest and the large size of the colony.
The displays that occur with the reassembly of the colony and the finding of mates, as well as those preceding copulation, are quite similar among the majority of species, but the accompanying vocalizations are more diverse. Various species have been described as trumpeting, croaking, cackling, and cooing; members of the genus Spheniscus are called jackass penguins for the braying sounds they make. The behaviour of experienced older birds is more elaborate and more effective than that of younger individuals. For example, Adélie penguins (Pygoscelis adeliae) may return to the reproductive colony from their third year onward but do not breed successfully until their fifth or sixth year.
Incubation of eggs is performed by both sexes in all species except the emperor penguin, in which it is done exclusively by the male, and it is begun immediately after egg laying. With the advent of incubation, the bustle and myriad cries that characterized mating give way to quiet and inactivity. Faulty incubation behaviour by inexperienced birds frequently results in the abandonment or breakage of eggs. The mortality rate (eggs and chicks) is very important at the egg stage, varying from year to year depending on climatic conditions, the percentage of young birds in the reproductive population, and the pressure of predation. In general, mortality (eggs and chicks) is from 40 to 80 percent of the eggs laid. In coastal colonies predators include, in order of importance: skuas, sheathbills, and the giant petrel. On the Australian, African, and South American continents, the nocturnal habits of certain penguins and the fact that they nest in burrows substantially limit predation, which is mostly by gulls and man.
Following egg laying, the female usually departs for the sea to feed, returning to relieve her mate after about 10 to 20 days. Thereafter, father and mother alternate in periods of a week or two. The female emperor penguin, however, must often walk 80 to 160 km (50 to 100 miles) from the colony to the sea and does not return until the end of the incubation period. During the 64-day incubation period, which extends through the height of the Antarctic winter, the male emperor penguin incubates the egg, holding it on his feet and living on stored fat reserves. During violent winter storms, members of the colony gather for mutual protection from wind and cold in tightly packed crowds called huddles.
Emergence from the shell takes 24 to 48 hours, during which the brooding parent is particularly irritable. The chick shows feeding behaviour immediately on hatching, receiving a regurgitated “soup” of crustaceans or fish by inserting its bill into the open mouth of the parent. During its early days the young bird is sheltered under the body of one of its parents, who take turns foraging and brooding. Growing larger, the young bird remains at a parent’s side, although the fledgling is able to maintain its body heat and move about alone. The chick then joins 100 or more of its contemporaries in a nursery group, or crèche, sometimes guarded by a few adults, while both its parents forage at sea. Upon returning with food, the parent calls its chick from the crèche and is able to distinguish it from other chicks (which frequently respond) by voice and appearance.
During the breeding season the number of “unemployed” adults in the colony increases with the addition of those who have lost eggs or chicks. In emperor penguin colonies, these unemployed birds often interfere with parents that have young and cause increased mortality. During the crèche stage the fuzzy down that has covered the chick since hatching is replaced by a coat of short stiff feathers, which are similar to those of the adult but usually somewhat different in colour. Once this molt is complete, the juvenile leaves the colony to seek its own food at sea.
The period of growth of the young bird from hatching to complete independence varies from two months, in the smallest species of the genus Eudyptula, to 51/2 months in the emperor and 12 to 14 months in the king penguin.
Adult penguins molt all of their feathers once a year following the breeding period. While in molt the bird is unable to enter the water and instead withdraws to a communal molting site usually situated in a sheltered area away from the colony. The duration of the molt varies from about two weeks in small species to more than a month in the larger ones.
The principal enemies of penguins at sea are the leopard seal and the killer whale (orca). Seals also take penguins near Australia, New Zealand, and other subantarctic regions.
Locomotion and orientation
Penguins are adapted for rapid locomotion in water, in which the wings, or flippers, are used for propulsion; the birds “fly” underwater. When moving at high speed, they frequently leave the water in leaps that may carry them a metre or more through the air; it is during this time that they breathe.
On land, penguins are much more awkward, even amusing, as they rock from side to side as they walk. Despite their short legs, however, penguins can run with surprising speed. Some, such as the northern rockhopper (Eudyptes moseleyi), the southern rockhopper (E. chrysocome), and Adélie penguins, move among rocks with agility, using the flippers for balance. On snow or ice, many penguins “toboggan,” sliding on the belly as they propel themselves with the feet and flippers. The flippers, along with the beak, are the prime weapons in defense and attack.
Scientists have long wondered how penguins are able to find their way back to their colonies from far out at sea, where currents may have carried them great distances. Also perplexing is how they are able to direct themselves correctly on land in the absence of clear-cut landmarks. Studies of penguins transported to the interior of Antarctica have found that they are able to find their way back to the ocean by using the sun as a directional aid. It is probable that the same means of orientation is used at sea. Upon approaching the coast they are able to recognize features of the shoreline and ocean bottom.
Food habits
The type of food utilized varies with the species, the geographic region, and the time of year. Most of the smaller southern penguins feed primarily upon krill, which attain high densities in the rich, well-oxygenated Antarctic waters. Cephalopods (squid and cuttlefish) and small fishes may form substantial fractions of the food, and in a few, such as the African penguin, fish is the basic element of the diet. The total weight of food consumed by a large penguin colony is prodigious, often exceeding several tons per day.
Form and function
The penguins are highly specialized for their flightless aquatic existence. The feet are located much farther back than those of other birds, with the result that the bird carries itself mostly upright; its walk can thus be described as plantigrade (i.e., on the soles). The sole comprises the whole foot instead of just the toes, as in other birds. The most notable characteristic of the group is the transformation of the forelimb into a paddle. This is accompanied by a body morphology particularly adapted to movement in a liquid medium. The thoracic (rib) cage is well developed, and the sternum bears a pronounced keel for the attachment of the pectoral muscles, which move the flippers. The flipper has the same skeletal base as the wing of flying birds but with its elements shortened and flattened, producing a relatively rigid limb covered with very short feathers—an ideal organ for rapid propulsion. The body plumage likewise consists of very short feathers, which minimize friction and turbulence. The density of the plumage and the layer of air that it retains provide almost complete insulation of the body.
Insulation of the bird’s body is particularly important for Antarctic species that live in water that is always below 0 °C (32 °F). The cooling power of seawater at −1.9 °C (28.6 °F) is equal to that of a temperature of −20 °C (−4 °F) with a wind of 110 km (70 miles) per hour. The skin is insulated by a layer of air trapped under the plumage, and the only bare skin in direct contact with the water is that of the feet. In the case of the emperor penguin on land, the feet are in almost constant contact with ice. The skin temperature is in the neighbourhood of 0 °C, and snow does not melt upon contact. This is possible because of remarkable anatomical arrangements in the lower limb, whereby closely adjacent arteries and veins form a system of heat exchange between opposing flows of blood. This arrangement permits cooled blood from the feet to absorb heat from outflowing blood, providing maximum economy of heat consistent with the functioning of the foot.
Like other seabirds, penguins have salt glands that enable them to ingest salt from seawater. Excess chloride is excreted in the form of a solution the concentration of which is greater than that of seawater. These glands are located above the eyes and are already functional in the young chick, which begins to consume food of marine origin from its first day of life.
Recent research has shown that the species most isolated geographically, such as the emperor penguin, can be subject to diseases. Some, such as the Adélie penguin, carry in their bodies trace amounts of pollutants, albeit in lower quantities than are found in many birds that live closer to humans.
Evidence from paleontology indicates that the penguins and the order Procellariiformes (albatrosses, shearwaters, and petrels) had a common origin. Both groups are represented by well-defined fossils dating to about 50 million years ago. The flightless sphenisciform line produced a number of distinctive side branches, all recognizably penguins, some giant in size. All of the fossil remains of penguins have been collected within the zone of the present-day distribution of the Sphenisciformes. Some apparently lived in warmer regions than do most of today’s penguins.
Phylogenetic analysis of living and fossil penguins shows that the group evolved a large body size early in its history. For example, two of the largest fossil penguins known—Icadyptes, which stood some 1.5 metres (about 5 feet) tall, and math, which stood about 1.8 metres (6 feet) tall—date to the Eocene Epoch (56 million to 33.9 million years ago). Living penguins make up a separate lineage characterized by smaller, highly aquatic species that began about 8 million years ago. The comparatively small size of living penguins is thus a geologically recent phenomenon that postdates the original radiation of giant penguins.
Additional Information
Penguins are a family of 17 to 19 species of birds that live primarily in the Southern Hemisphere. They include the tiny blue penguins of Australia and New Zealand, the majestic emperor penguins of Antarctica and king penguins found on many sub- Antarctic islands, the endangered African penguin and the Galápagos penguin—the only penguin to be found north of the equator.
Though they are birds, penguins have flippers instead of wings. They cannot fly and on land they waddle walking upright—though when snow conditions are right they will slide on their bellies. In the water they are expert swimmers and divers, and some species can reach speeds of up to 15 miles per hour. The penguin’s distinctive coloring—black body with white belly—helps camouflage the bird in the water as it searches for meals of small shrimp, fish, crabs and squid.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2581 2025-09-05 18:12:25
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2381) Niagra Falls
Gist
What makes Niagara Falls a natural wonder? Statistically speaking, Niagara Falls is recognized as having the greatest flow rate of any waterfall throughout the globe. The falls have more than six million cubic feet (168,000 cubic meters) flow over the top of the falls every minute during the peak season.
Niagara Falls is a group of three waterfalls—American, Bridal Veil, and Horseshoe Falls—located on the Niagara River at the border between Ontario, Canada, and New York, USA. Known for the immense flow of water and spectacular views, the falls have the highest flow rate of any waterfall globally. The site is a popular tourist attraction, a source of hydroelectric power, and home to Niagara Falls State Park, the oldest state park in the United States.
The falls consist of the American Falls, the Bridal Veil Falls, and the much larger Horseshoe Falls.
Summary
Niagara Falls[a] is a group of three waterfalls at the southern end of Niagara Gorge, spanning the border between the province of Ontario in Canada and the state of New York in the United States. The largest of the three is Horseshoe Falls, which straddles the international border of the two countries. It is also known as the Canadian Falls. The smaller American Falls and Bridal Veil Falls lie within the United States. Bridal Veil Falls is separated from Horseshoe Falls by Goat Island and from American Falls by Luna Island, with both islands situated in New York.
Formed by the Niagara River, which drains Lake Erie into Lake Ontario, the combined falls have the highest flow rate of any waterfall in North America that has a vertical drop of more than 50 m (164 ft). During peak daytime tourist hours, more than 168,000 {m}^{3} (5.9 million cu ft) of water goes over the crest of the falls every minute. Horseshoe Falls is the most powerful waterfall in North America, as measured by flow rate. Niagara Falls is famed for its beauty and is a valuable source of hydroelectric power. Balancing recreational, commercial, and industrial uses has been a challenge for the stewards of the falls since the 19th century.
Niagara Falls is 27 km (17 mi) northwest of Buffalo, New York, and 69 km (43 mi) southeast of Toronto, between the twin cities of Niagara Falls, Ontario, and Niagara Falls, New York. Niagara Falls was formed when glaciers receded at the end of the Wisconsin glaciation (the last ice age), and water from the newly formed Great Lakes carved a path over and through the Niagara Escarpment en route to the Atlantic Ocean.
Details
Niagara Falls, waterfall on the Niagara River in northeastern North America, one of the continent’s most famous spectacles. The falls lie on the border between Ontario, Canada, and New York state, U.S. For many decades the falls were an attraction for honeymooners and for such stunts as walking over the falls on a tightrope or going over them in a barrel. Increasingly, however, the appeal of the site has become its beauty and uniqueness as a physical phenomenon.
The falls are in two principal parts, separated by Goat Island. The larger division, adjoining the left, or Canadian, bank, is Horseshoe Falls; its height is 188 feet (57 metres), and the length of its curving crest line is about 2,200 feet (670 metres). The American Falls, adjoining the right bank, are 190 feet (58 metres) high and 1,060 feet (320 metres) across.
The formation of the Niagara gorge (downriver) and the maintenance of the falls as a cataract depend upon peculiar geologic conditions. The rock strata from the Silurian Period (about 444 to 419 million years ago) in the Niagara gorge are nearly horizontal, dipping southward only about 20 feet per mile (almost 4 metres per km). An upper layer of hard dolomite is underlain by softer layers of shale. Water exerts hydrostatic pressure and only slowly dissolves the dolomite after infiltrating its joints. Dolomite blocks fall away as water from above infiltrates and rapidly erodes the shale at the falls itself. The disposition of the rock strata provides the conditions for keeping the water constantly falling vertically from an overhanging ledge during a long period of recession (movement upstream) of the cataract. As blocks of dolomite are undercut, they fall off and are rapidly destroyed by the falling water, further facilitating the retreat of the falls and the maintenance of a vertical cataract.
The water flowing over the falls is free of sediment, and its clearness contributes to the beauty of the cataract. In recognition of the importance of the waterfall as a great natural spectacle, the province of Ontario and the state of New York retained or acquired title to the adjacent lands and converted them into public parks.
The very large diversion of water above the falls for hydroelectric power purposes has lessened the rate of erosion. Elaborate control works upstream from the falls have maintained an even distribution of flow across both the U.S. and Canadian cataracts, thereby preserving the curtains of the waterfalls. A large part of the great river above the falls is diverted and disappears into four great tunnels for use in the power plants downstream. Because of concern over the possibility of major rockfalls, water was diverted from the American Falls in 1969, and some cementing of the bedrock was done; an extensive boring and sampling program was also carried out. River flow was returned to the American Falls in November of that year, and it was decided that safety measures for the viewing public should be implemented and that measures to stem natural processes were both too expensive and undesirable.
Excellent views of the falls are obtained from Queen Victoria Park on the Canadian side; from Prospect Point on the U.S. side at the edge of the American Falls; and from Rainbow Bridge, which spans the Niagara gorge about 1,000 feet (300 metres) downstream from Prospect Point. Visitors may cross from the U.S. shore to Goat Island by footbridge and may take an elevator to the foot of the falls and visit the Cave of the Winds behind the curtain of falling water. The Horseshoe Falls, which carry about 90 percent of the river’s discharge, receded upstream at an average rate of about 5.5 feet (1.7 metres) per year in 1842–1905. Thereafter, control works and the diversion of water decreased the erosion rate, which is presently so slow at the American Falls that large blocks of dolomite accumulate at the base of the falls, threatening to turn it into rapids.
Additional Information
Situated between the state of New York and the province of Ontario, Niagara Falls is one of the most spectacular natural wonders in North America. This series of waterfallsis on the Niagara River, which flows between the United States and Canada from Lake Erie to Lake Ontario. The falls are about 17 miles (27 kilometers) northwest of Buffalo, New York. The industrial city and tourist center of Niagara Falls, New York, is adjacent to the American side of the falls. Niagara Falls, Ontario, Canada, is across the river.
The falls are divided into two parts by Goat Island. The larger portion, on the southwest side, is the Canadian falls, known as the Horseshoe Falls. It measures 2,600 feet (790 meters) along its curve and drops 162 feet (49 meters). The smaller American Falls is northeast of Goat Island. It is 1,000 feet (305 meters) across and drops about 167 feet (51 meters). Between the American Falls and Goat Island are the small Luna Island and the small Luna, or Bridal Veil, Falls.
Just before flowing over the ledge, the American stream is only about 3 1/2 feet (1 meter) deep. The Canadian stream is about 20 feet (6 meters) deep and carries some 95 percent of the Niagara River’s water.
Every minute about 12,000,000 cubic feet (340,000 cubic meters), or 379,000 tons, of water pours in torrents over the cliffs of the falls of Niagara. As the water plunges from the brink of the falls, it fills the air with a silvery mist, which under the sunlight displays many rainbows. The plunging water also sends out a never-ending roar as it strikes the bottom. For this reason the Haudenosaunee (Iroquois) people called the falls Niagara, meaning “thunder of waters.”
The plunging water has worn the lower rocks away so that there are caves behind the sheets of water of both falls. Sightseers may enter the Cave of the Winds at the foot of the American Falls and get an unusual view. The Horseshoe Falls have carved a plunge basin 192 feet (59 meters) deep.
Both the United States and Canadian governments have built parks, viewing platforms, paths, and highways. The Niagara Reservation State Park was established in 1885 and is New York’s oldest state park. It includes an observation tower, elevators that descend into the gorge at the base of the American Falls, and boat trips into the waters at the base of the Horseshoe Falls.
The park area has long been a tourist site and a favorite spot for couples to spend their honeymoons. At night colored lights illuminate the falls.
Almost all the drainage from four of the Great Lakes pours over the crest of Niagara Falls. This tremendous volume of water is used to generate power in six hydroelectric plants. They develop a maximum of about 5 3/4 million horsepower, some 55 percent on the American side and about 45 percent on the Canadian. The plants draw water from the river above the falls through canals. Near each plant the water drops through penstocks to powerhouses on the Niagara River below the falls. There it turns turbine generators.
The control of Niagara Falls between the United States and Canada has long offered the world an example of international cooperation. A treaty in 1910 and later agreements fixed the amounts of water that could be diverted. An international Niagara Control Board was established in 1923.
In 1950 the two countries signed a new treaty that specified the minimum flow to be maintained over the falls. This treaty made possible greater hydroelectric development. It provides that 100,000 cubic feet (2,830 cubic meters) per second of water must flow over the falls during the tourist season in the daytime and 50,000 (1,415) at night and during the off-tourist season in the daytime. The remainder is equally divided between Canada and the United States. An average of 202,300 cubic feet (5,729 cubic meters) per second flows over the falls.
Between 1954 and 1958 the United States and Canada completed the Niagara Remedial Works Project. This enormous operation checked erosion with a gated control structure, excavations, and fills.
The Hydro-Electric Power Commission of Ontario completed the Sir Adam Beck-Niagara Generating Station No. 1 in 1925 and No. 2 in 1958. The combined capacity of the plants is 1,443,000 kilowatts.
In 1957 the United States Congress approved the construction of the Niagara Power Project by the Power Authority of the State of New York. It has a capacity of 2,190,000 kilowatts. The first electric current from the project was delivered in 1961.
The falls of Niagara are about 25,000 years old. The hard rock (Lockport dolomite) at the brink of the falls is much older. It was made on the bed of an inland sea in the Silurian period (about 443 million to 419 million years ago). Gradually the limy sediment hardened to stone—either limestone or dolomite, a limestone with magnesium.
Later the Niagara region was raised in a widespread uplift centered in Michigan. Streams wore down the land. The layer of tough rock, however, resisted erosion. The edge of the deposit formed a great cliff—the Niagara escarpment. It runs west from Rochester, New York, between Lakes Erie and Ontario, then swings northward through the province of Ontario. It is capped by hard Niagara limestone or Lockport dolomite.
Glaciers covered the Niagara region during the Ice Age that took place during the Pleistocene Epoch (about 2.6 million to 11,700 years ago). As the last glacier retreated, it left Lake Erie at its southern edge. Water from the lake began to spill over the Niagara escarpment into the Ontario basin below, just south of where Queenston and Lewiston now stand.
The new falls did not wear away the dolomite caprock as fast as it churned away the softer rock below. From time to time blocks of the undermined caprock broke off. The falls worked back toward Lake Erie, forming a steep-walled gorge.
Niagara’s rate of cutting has changed many times. It started slowly, for at first the river drained Lake Erie only. Lakes Superior, Michigan, and Huron had a northerly outlet. The drainage changed as glaciers retreated. Water from all four lakes then poured over the falls. When the river spread to the point where the famous whirlpool now is, it reached an ancient valley that had cut into the dolomite from the west. Later the valley filled with glacial debris. The river wore away the soft material, forming the 60-acre (24- hectare) basin.
Louis Hennepin, a priest who accompanied the French explorer René-Robert Cavelier, sieur de La Salle, was the first European to view Niagara Falls, in 1678. The site was of strategic use to the British and French in the struggle to control the Great Lakes. The British built Fort Schlosser there in 1761.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2582 2025-09-06 22:44:37
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2382) Congo River
Gist
The deepest river in the world is the Congo River in Africa, with recorded depths reaching at least 220 meters (720 feet). Recognized by Guinness World Records, its maximum depth surpasses that of any other river and was confirmed by scientists in 2008.
Summary
The basin of the deepest river in the world provides food, water, medicines and transportation to about 75 million people, according to Yale University’s Global Forest Atlas.
1 . Where does the Congo river flow through?
The Congo is the deepest river in the world. Its headwaters are in the north-east of Zambia, between Lake Tanganyika and Lake Nyasa (Malawi), 1760 metres above sea level; it flows into the Atlantic Ocean.
The Congo river flows through the Republic of the Congo, the Democratic Republic of the Congo, the Central African Republic, western Zambia, northern Angola, Cameroon and Tanzania. The lower course of the Congo river has large gorges and falls, which makes it one of the most dangerous rivers.
2 . How deep and how long is the Congo river?
The Congo river, formerly known as the Zaire, is the deepest river in the world: at some points the water can be up to 220 metres deep; the basin has a surface area of 3,457,000 square kilometres.
The Congo river is 4,700 kilometres long, which makes it the second longest river in Africa, the tenth longest river in the world and the second largest river by discharge volume in the world, only after the Amazon.
3 . What are the environmental threats in the Congo river basin?
One of the main threats in the Congo river basin is deforestation to accommodate modern agricultural practices; moreover, industrial logging accelerates the rate of deforestation.
The other major threat is food demand due to rapid population growth in the region, which leads to excessive hunting of wild animals such as bats, monkeys, rats and snakes.
Summary
he Congo River, formerly also known as the Zaire River, is the second-longest river in Africa, shorter only than the Nile, as well as the third-largest river in the world by discharge volume, following the Amazon and Ganges–Brahmaputra rivers. It is the world's deepest recorded river, with measured depths of around 220 m (720 ft). The Congo–Lualaba–Luvua–Luapula–Chambeshi River system has an overall length of 4,700 km (2,900 mi), which makes it the world's ninth-longest river. The Chambeshi is a tributary of the Lualaba River, and Lualaba is the name of the Congo River upstream of Boyoma Falls, extending for 1,800 km (1,100 mi).
Measured along with the Lualaba, the main tributary, the Congo River has a total length of 4,370 km (2,720 mi). It is the only major river to cross the equator twice. The Congo Basin has a total area of about 4,000,000 sq km (1,500,000 sq mi), or 13% of the entire African landmass.
Name
The name Congo/Kongo originates from the Kingdom of Kongo once located on the southern bank of the river. The kingdom in turn was named after the indigenous Bantu Kongo people, known in the 17th century as "Esikongo". South of the Kingdom of Kongo proper lay the similarly named Kakongo kingdom, mentioned in 1535. Abraham Ortelius labelled "Manicongo" as the city at the mouth of the river in his world map of 1564. The tribal names in Kongo possibly derive from a word for a public gathering or tribal assembly. The modern name of the Kongo people or Bakongo was introduced in the early 20th century.
The name Zaire is from a Portuguese adaptation of a Kikongo word, nzere ("river"), a truncation of nzadi o nzere ("river swallowing rivers"). The river was known as Zaire during the 16th and 17th centuries; Congo seems to have replaced Zaire gradually in English usage during the 18th century, and Congo is the preferred English name in 19th-century literature, although references to Zahir or Zaire as the name used by the inhabitants remained common. The Democratic Republic of the Congo and the Republic of the Congo are named after it, as was the previous Republic of the Congo which had gained independence in 1960 from the Belgian Congo. The Republic of Zaire during 1971–1997 was also named after the river's name in French and Portuguese.
Details
Congo River, river in west-central Africa. With a length of 2,900 miles (4,700 km), it is the continent’s second longest river, after the Nile. It rises in the highlands of northeastern Zambia between Lakes Tanganyika and Nyasa (Malawi) as the Chambeshi River at an elevation of 5,760 feet (1,760 metres) above sea level and at a distance of about 430 miles (700 km) from the Indian Ocean. Its course then takes the form of a giant counterclockwise arc, flowing to the northwest, west, and southwest before draining into the Atlantic Ocean at Banana (Banane) in the Democratic Republic of the Congo. Its drainage basin, covering an area of 1,335,000 square miles (3,457,000 square km), takes in almost the entire territory of that country, as well as most of the Republic of the Congo, the Central African Republic, eastern Zambia, and northern Angola and parts of Cameroon and Tanzania.
With its many tributaries, the Congo forms the continent’s largest network of navigable waterways. Navigability, however, is limited by an insurmountable obstacle: a series of 32 cataracts over the river’s lower course, including the famous Inga Falls. These cataracts render the Congo unnavigable between the seaport of Matadi, at the head of the Congo estuary, and Malebo Pool, a lakelike expansion of the river. It was on opposite banks of Malebo Pool—which represents the point of departure of inland navigation—that the capitals of the former states of the French Congo and the Belgian Congo were founded: on the left bank Kinshasa (formerly Léopoldville), now the capital of the Democratic Republic of the Congo, and on the right bank Brazzaville, now the capital of the Republic of the Congo.
The Amazon and the Congo are the two great rivers of the world that flow out of equatorial zones where heavy rainfall occurs throughout all or almost all of the year. Upstream from Malebo Pool, the Congo basin receives an average of about 60 inches (1,500 mm) of rain a year, of which more than one-fourth is discharged into the Atlantic. The drainage basin of the Congo is, however, only about half the size of that of the Amazon, and the Congo’s rate of flow—1,450,000 cubic feet (41,000 cubic metres) per second at its mouth—is considerably less than the Amazon’s flow of more than 6,180,000 cubic feet (175,000 cubic metres) per second.
While the Chambeshi River, as the remotest source, may form the Congo’s original main stream in terms of the river’s length, it is another tributary—the Lualaba, which rises near Musofi in southeastern Democratic Republic of the Congo—that carries the greatest quantity of water and thus may be considered as forming the Congo’s original main stream in terms of water volume.
When the river first became known to Europeans at the end of the 15th century, they called it the Zaire, a corruption of a word that is variously given as nzari, nzali, njali, nzaddi, and niadi and that simply means “river” in local African languages. It was only in the early years of the 18th century that the river was first called the “Rio Congo,” a name taken from the kingdom of Kongo that had been situated along the lower course of the river. During the period (1971–97) when the Democratic Republic of the Congo was called Zaire, the government also renamed the river the Zaire. Even during that time, however, the river continued to be known throughout the world as the Congo. To the literary-minded the river is evocative of the famous 1902 short story “Heart of Darkness” by Joseph Conrad. His book conjured up an atmosphere of foreboding, treachery, greed, and exploitation. Today, however, the Congo appears as the key to the economic development of the central African interior.
Physical features:
Physiography
The expression “Congo basin,” strictly speaking, refers to the hydrographic basin. This is not only vast but is also covered with a dense and ramified network of tributaries, subtributaries, and small rivers—with the exception of the sandy plateaus in the southwest.
The Congo basin is the most clearly distinguished of the various geographic depressions situated between the Sahara to the north, the Atlantic Ocean to the south and west, and the region of the East African lakes to the east. In this basin, a fan-shaped web of tributaries flows downward along concentric slopes that range from 900 to 1,500 feet (275 to 460 metres) in elevation and that enclose a central depression. The basin itself stretches for more than 1,200 miles (1,900 km) from north to south (from the Congo–Lake Chad watershed to the interior plateaus of Angola) and also measures about 1,200 miles from the Atlantic in the west to the Nile-Congo watershed in the east.
The central part of the Congo basin—often called the cuvette (literally “saucer” or “shallow bowl”)—is an immense depression containing Quaternary alluvial deposits that rest on thick sediments of continental origin, consisting principally of sands and sandstones. These underlying sediments form outcrops in valley floors at the eastern edge of the cuvette. The filling of the cuvette, however, began much earlier. Boreholes have revealed that since late Precambrian times (i.e., since at least 570 million years ago) considerable sediment has accumulated, derived from the erosion of formations situated around the periphery of the cuvette. The arrangement of surface relief, thick depositional strata, and substratum in amphitheatre-like fashion around the main Congo channel, which has been uniform across time, is evidence of a persistent tendency to subsidence in this part of the continent. This subsidence is accompanied by uplifting on the edges of the cuvette, principally on its eastern side—which has also been influenced by the formation of the Western Rift Valley.
From its sources to its mouth, the Congo River system has three contrasting sections—the upper Congo, middle Congo, and lower Congo. The upper reaches are characterized by three features—confluences, lakes, and waterfalls or rapids. To begin with, several streams of approximately equal size unite to form the river. In a little more than 60 miles (100 km), the upper Lualaba joins the Luvua and then the Lukuga. Each stream for part of its course undergoes at least a lacustrine type of expansion, even when it does not form a lake. Thus, Lake Upemba occurs on the upper Lualaba; Lakes Bangweulu and Mweru occur on the Chambeshi–Luapula–Luvua system; and Lake Tanganyika, which is fed by the Ruzizi (flowing from Lake Kivu) and by the Malagarasi, itself flows into the Lukuga. Rapids occur not only along the headstreams but also in several places along the course of the main stream. Navigation thus is possible only along sections of the upper Congo by vessels of low tonnage. Even so, these stretches are in danger of being overgrown by aquatic vegetation, particularly water hyacinths.
Kisangani (formerly Stanleyville)—located just downstream of the Boyoma Falls, a series of seven cataracts—marks the real beginning, upriver, of the navigable Congo. This central part of the river flows steadily for more than 1,000 miles (1,600 km) to within 22 miles (35 km) of Kinshasa. Its course at first is narrow but soon grows wider, after which many islands occur in midstream. This change in the character of the river corresponds to its entry into its alluvial plain. From that point onward, with the exception of a few rare narrow sections, the Congo divides into several arms, separated by strings of islands. It increases from a width of more than 3.5 miles (5.5 km) downstream from Isangi (where the Lomami enters the Congo) to a width of 5 to 7 miles (8 to 11 km) and on occasion—for example, at the mouth of the Mongala—to 8 miles (13 km). Beyond the natural levees (formed by silt deposits) occurring on either bank, some areas are subjected to extensive flooding that increases the river’s bounds still further. It is not always easy to distinguish such areas from the “rain swamps” in regions lying between rivers. The middle course of the Congo ends in a narrow section called the Chenal (“Channel”), or Couloir (“Corridor”). Between banks no more than half a mile to a mile wide, the riverbed deepens and the current becomes rapid, flowing through a valley that cuts down several hundreds of yards deep into the soft sandstone bedrock of the Batéké Plateau. Along this central reach the Congo receives its principal tributaries, primarily the Ubangi and the Sangha on the right bank and the Kwa on the left bank. An enormous increase in the average rate of flow results, rising from less than 250,000 cubic feet (7,000 cubic metres) a second at Kisangani to nearly its maximum flow at Kinshasa.
Upon leaving the Chenal, the Congo divides into two branches, forming Malebo Pool, a vast lacustrine area about 15 by 17 miles (24 by 27 km), which marks the end of the middle Congo. Immediately downstream occur the first waterfalls of the final section of the river’s course. Cataracts and rapids are grouped into two series, separated by a fairly calm central reach, in which the elevation drops from a little less than 900 feet (275 metres) to a few yards above sea level. The Congo’s estuary begins at Matadi, downstream from the rapids that close off the interior Congo; 83 miles (134 km) in length, it forms the border between Angola and the Democratic Republic of the Congo. At first the estuary is narrow—less than half a mile to about a mile and a half in width—with a central channel 65 to 80 feet (20 to 24 metres) deep, but it widens downstream of Boma. There the river, obstructed by islands, divides into several arms, and in some places the depth does not exceed 20 to 25 feet (6 to 7.5 metres), which makes dredging necessary to allow oceangoing vessels to reach Matadi. Beyond the estuary’s mouth, the course of the Congo continues offshore as a deep underwater canyon that extends for a distance of about 125 miles (200 km).
Hydrology of the Congo River
The Congo has a regular flow, which is fed by rains throughout the year. At Kinshasa the flow has for many years remained between the high level of 2,310,000 cubic feet (65,000 cubic metres) per second, recorded during the flood of 1908, and the low level of 756,000 cubic feet (21,000 cubic metres) per second, recorded in 1905. During the unusual flood of 1962, however, by far the highest for a century, the flow probably exceeded 2,600,000 cubic feet (73,000 cubic metres) per second.
At Brazzaville and Kinshasa, the river’s regime is characterized by a main maximum at the end of the year and a secondary maximum in May, as well as by a major low level during July and a secondary low level during March and April. In reality, the downstream regime of the Congo represents climatic influence extending over 20° of latitude on both sides of the Equator a distance of some 1,400 miles (2,250 km). Each tributary in its course modifies the level of the main stream. Thus, for example, the low level in July at Malebo Pool results from two factors: a drought that occurs for several months in the southern part of the basin at that time, as well as a delay before the floods of the Ubangi tributary flowing down from the north arrive, which does not happen before August. The Congo basin is so vast that no single meteorologic circumstance is capable of disturbing the slow movement of the waters’ rise and fall. The annual fluctuations may alter drastically, however, when floodwaters from different tributaries that normally coincide with each other arrive at different times.
Lake Tanganyika, apart from brief seiches caused by wind drift and sudden changes in atmospheric pressure, may experience considerable variations in its water level from year to year. In 1960, for example, its waters flooded parts of Kalemi, Democratic Republic of the Congo, and Bujumbura, Burundi. A series of particularly rainy years followed by a blocking of the outlet by floating vegetation may explain this phenomenon.
Climate
Typical climate in regions through which the Congo flows is that of Yangambi, a town situated on the river’s right bank slightly north of the Equator and a little downstream of Kisangani. Humidity is high throughout the year, and annual rainfall amounts to 67 inches (1,700 mm) and occurs fairly regularly; even in the driest month the rainfall totals more than 3 inches (76 mm). Temperatures are also uniformly high throughout the year, and there is little diurnal variability. The average temperature at Yangambi is in the mid-70s F (mid-20s C).
From the pluviometric equator (an imaginary east-west line indicating the region of heaviest rainfall), which is situated slightly to the north of the geographic equator, the amount of rainfall decreases regularly in proportion to latitude. The northernmost points of the basin, situated in the Central African Republic, receive only from 8 to 16 inches (200 to 400 mm) less during the course of a year than points near the Equator. The dry season, however, lasts for four or five months, and there is only one annual rainfall maximum, which occurs in summer. In the far southern part of the basin, at a latitude of 12° S, the climate becomes definitely Sudanic in character, with marked dry and wet seasons of approximately equal length and with rainfall of about 49 inches (1,250 mm) a year.
Plant life
The Congo basin is home to the second largest rainforest in the world. The equatorial climate that prevails over a significant part of the Congo basin is coextensive with a dense evergreen forest. The Congolese forest spreads out over the central depression, extending continuously from about 4° N to about 5° S; it is interrupted only by clearings, many of which have a natural origin. The forest region is bordered on either side by belts of savanna (grassy parkland). The forest and savanna often meet imperceptibly, blending together in a mosaic pattern; more rarely, strips of forest invade the grassland. Farther away from the Equator, and to the extent that the Sudanic features of the climate become evident, the wooded savanna region, with its thin deciduous forest, is progressively reached.
As it courses through the solid mass of the Congolese forest, the Congo and its tributaries are bordered by discontinuous grassy strips. Meadows of Echinochloa (a type of grass), papyrus, and Cyperaceae (sedge) occupy abandoned river channels, fringe the banks, or, behind a curtain of forest, blanket the depressions in the centre of the islands, They also spring up on sandbanks, as well as on the downstream ends of islands that are fertilized by the floods. A shrub, Alchornea, frequently marks the transition to the high forest that grows on the levees behind the banks.
Animal life
The animal life of the Congo basin is identified to a certain extent with that of the equatorial forest, which is sharply distinct from the wildlife of the savannas. Within this equatorial domain, the Congo and its principal tributaries form a separate ecological milieu. The animal population of the great waterways often has fewer affinities with the neighbouring marshes or the forests on dry land than it has with other river systems, whether of the coastal region or the savannas.
Numerous species of fish live in the waters of the Congo; more than 230 have been identified in Malebo Pool and the waters that flow into it alone. The riverine swamps, which often dry up at low water, are inhabited by lungfish, which survive the dry periods buried and encysted in cocoons of mucus. In the wooded marshlands, where the water is the colour of black tea, the black catfish there assume the colour of their environment. The wildlife of the marshes and that of the little parallel streams do not mix with the wildlife of the river itself.
The waters of the Congo contain various kinds of reptiles, of which crocodiles are the most striking species. Semiaquatic tortoises are also found, as are several species of water snakes.
The forest birdlife constitutes, together with the birdlife of the East African mountains, the most specifically indigenous birdlife found on the African continent. In the Congo region more than 265 species typical of the equatorial forest have been recorded. Occasionally or seasonally, however, nontypical birds may be observed. Seabirds, such as the sea swallow, fly upstream from the ocean. Migratory birds from Europe, including the blongios heron and the Ixobrychus minutus (little bittern), pass through the region. Species with a wide distribution within Africa, such as the Egyptian duck, also have been sighted. Ducks, herons, storks, and pelicans are abundantly represented.
Aquatic mammals are rare, consisting of the hippopotamus, two species of otters, and the manatee. The manatee (sea cow), which lives entirely in the water, has been officially identified only on the Sangha tributary but appears to have given rise to some curious legends on the lower Congo, including its association with a creature called Mami Wata (a kind of siren), stories of which were carried by African slaves to the Americas.
The people and the economy:
Life of the river peoples
Three types of environments are found, either juxtaposed or in succession, along the river and its tributaries: the narrower sections, bordered by firm ground; the wider stretches, dotted with islands and accompanied by backwaters; and the zones where flooding occurs or where there are extensive marshes.
Almost all the river peoples engage in fishing. Along the narrow sections, where rapids often occur, fishing is only of interest to a small number of villages. The Enya (Wagenia) of Boyoma Falls and the Manyanga living downstream from Malebo Pool attach fish traps to stakes or to dams built in the rapids themselves. Fishing of a very different nature, notably by poison, is conducted in the marshy areas, where the population is more extensive than might be imagined. Among these peoples are the Ngombe—“water people”—who inhabit the Itimbiri-Ngiri and the triangle formed by the Congo and the Ubangi. Other fisherfolk of the marshes dwell in the lagoons and the flooded forests of the region where the confluence of the Congo and the Alima, Likouala, and Sangha occurs.
Despite unfavourable conditions, all these peoples are also cultivators. They raise dikes, often of monumental size, to plant cassava (manioc) on the land thus sheltered from flooding. Other minor crops, such as sweet potatoes, bananas, and yams, are also found. The Congo basin has the continent’s most important timber resources, but the timber industry is developing slowly, mainly because the interior is so inaccessible and because the cost of transporting timber to the coast is so high.
Few modes of existence have undergone such profound changes as a result of contact with the modern world as has that of the river’s fisherfolk. The growth of the towns on the banks of Malebo Pool as well as the taste of urban dwellers for river fish have served to stimulate fishing by tying it to a cash economy. It is not just a question of villagers smoking fish that they sell to passing traders. Increasingly numerous fishing crews sail up the Congo, the Ubangi, and the Kasai, well above their confluences, to fish in the shallows.
Transportation
The Congo is an important navigational system in Africa. Within the territorial limits of the Democratic Republic of the Congo alone, there are some 8,700 miles (14,000 km) of navigable waterway. Of this total, 650 miles (1,050 km) are accessible at all seasons to barges with capacities between 800 and 1,100 tons, depending upon the height of the water. The amount of goods transported by water—consisting mainly of agricultural produce, wood, minerals, and fuel—is very modest in comparison with the traffic on European rivers (for example, the commercial traffic from the port of Kinshasa does not reach a million tons), but river transport remains essential for communications with regions that are inaccessible by road, especially in the cuvette. The three principal routes, all of which converge on the downstream terminus at Kinshasa on the Malebo Pool, run from Kisangani, from Ilebo (formerly Port-Francqui) on the Kasai, and from Bangui on the Ubangi. River transport, however, falls short of the role it could play in development. It has actually declined since the states of the Congo basin became independent in 1960, because of serious problems with aging equipment, a lack of maintenance of the infrastructure, and the poor functioning of the public waterway agencies. In the Democratic Republic of the Congo only the section from Ilebo to Kinshasa is still important, because it constitutes the river link (the other link being a railway between Kinshasa and Matadi) used to transport the copper production of Katanga to the coast.
This network has fostered economic development in inland areas, far from the coast. Varied activities include the production of palm oil on the banks of the Kwilu, centred on the port of Kikwit, and the establishment of plantations of robusta coffee in the Kisangani area.
Before such developments could be undertaken, however, it was necessary to overcome the barrier to the sea formed by the Congo’s lower course. That feat was accomplished in 1898 with the opening of the railway between Matadi and Léopoldville (now Kinshasa) and in 1934 by the completion of the Congo-Ocean rail line on the right bank between Brazzaville and Pointe-Noire.
While the river system facilitates navigation, it also hinders land transportation. Only a small number of bridges cross the Congo and its tributaries. The Kongolo rail-and-road bridge over the Lualaba was reconstructed in 1968, and a bridge over the Congo at Matadi was opened in 1983. Numerous projects to improve the situation nevertheless exist, notably a link between Kinshasa and Brazzaville. This project has long been under discussion, although to financial obstacles are added difficulties caused by political dissension. Several times since the two countries gained independence in 1960, dissension has interrupted the ferry traffic between the two capitals.
Power
It has been estimated that the hydroelectric potential of the Congo basin amounts to about one-sixth of the known world resources, but only a fraction of this potential has been harnessed. The single site of Inga, just upriver from Matadi, has a power potential estimated at more than 30,000 megawatts. Two hydroelectric projects, called Inga I and Inga II, have been completed there since the independence of the Democratic Republic of the Congo. Further development of the region’s hydropower potential, as outlined in the ambitious “Grand Inga” scheme, would create one of the world’s largest hydroelectric power systems.
Study and exploration
The question of the source of the Congo confronted European explorers from the time that the Portuguese navigator Diogo Cão encountered the river’s mouth in 1482, which he believed to be a strait providing access to the realm of the mythical Prester John, a Christian priest-king. It is virtually certain that, well before the Welsh explorer Henry Morton Stanley arrived in 1877, some 17th-century Capuchin missionaries reached the shores of Malebo Pool. This exploit, however, was not followed up, even by the amply supplied expedition led by James Kingston Tuckey, which was sent out by the British Admiralty in 1816 but was decimated and had to retrace its footsteps even before it had surmounted the cataracts. Preposterous hypotheses about the river continued to be entertained, connecting, for example, the upper Niger to the Congo or maintaining that the Congo and the Nile both flowed from a single great lake in the heart of Africa.
Even after the European discovery of Lake Tanganyika by the British explorers Richard Burton and John Speke (1858), then of the Lualaba (1867) and of Lake Bangweulu (1868) by the Scottish explorer David Livingstone, uncertainty remained—uncertainty that Stanley was to dissipate in the course of his famous expedition in 1876 and 1877 that took him by water from the Lualaba to the Congo’s mouth over a period of nine months. In the interior of the Congo basin and above all on the right bank, the final blank spaces on the map could not be filled in until about 1890, when the exploration of the upper course of the Ubangi was completed.
Additional Information
The Congo River (also known as Zaire River) is the largest river in Africa. Its overall length of 4,700 km (2,922 miles) makes it the second longest in Africa (after the Nile). The river and its tributaries flow through the second largest rain forest area in the world, second only to the Amazon Rainforest in South America.
The river also has the second-largest flow in the world, behind the Amazon, and the second-largest watershed of any river, again trailing the Amazon. Its watershed is a little larger than that of the Mississippi River. Because large parts of the river basin sit north and south of the equator, its flow is steady, as there is always at least one river having a rainy season. The Congo gets its name from the old Kingdom of Kongo which was at the mouth of the river. The Democratic Republic of the Congo and the Republic of the Congo, both countries sitting along the river's banks, are named after it. From 1971 to 1997, the Democratic Republic of the Congo was called Zaire and its government called the river the Zaire River.
The sources of the Congo are in the Highlands and mountains of the East African Rift, as well as Lake Tanganyika and Lake Mweru, which feed the Lualaba River. This then becomes the Congo below Boyoma Falls. The Chambeshi River in Zambia is usually taken as the source of the Congo because of the accepted practice worldwide of using the longest tributary, as with the Nile River.
The Congo flows mostly west from Kisangani just below the falls, then slowly bends southwest, passing by Mbandaka, joining with the Ubangi River, and running into the Pool Malebo (Stanley Pool). Kinshasa (formerly Léopoldville) and Brazzaville are on opposite sides of the river at the Pool, where the river narrows and falls through a few cataracts in deep canyons (collectively known as the Livingstone Falls), running by Matadi and Boma, and into the sea at the small town of Muanda.
History of exploration
The mouth of the Congo was visited by Europeans in 1482, by the Portuguese Diogo Cão, and in 1817, by a British exploration under James Kingston Tuckey that went up the river as far as Isangila. Henry Morton Stanley was the first European to travel along the whole river.
Economic importance
Although the Livingstone Falls stop ships coming in from the sea, almost all of the Congo is navigable in parts, especially between Kinshasa and Kisangani. Railways cross the three major falls that interrupt navigation, and much of the trade of central Africa passes along the river. Goods include copper, palm oil, sugar, coffee, and cotton. The river can also be valuable for hydroelectric power, and Inga Dams below Pool Malebo have been built.
In February of 2005, South Africa's state owned power company, Eskom, said that they had a proposal to increase the amount of electric power that the Inga can make through improvements and the building of a new hydroelectric dam. The project would bring the highest output of the dam to 40 GW, twice that of China's Three Gorges Dam.
Geological history
In the Mesozoic period before the continental drift opened the South Atlantic Ocean, the Congo was the upper part of a river about 12,000 km (7,500 miles) long that flowed west across the parts of Gondwanaland, now called Africa and South America.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2583 2025-09-07 21:28:45
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2383) Ostrich
Gist
Ostriches have special features including being the world's largest and fastest flightless bird, with powerful two-toed legs, largest eyes of any land animal, and unique two-toed feet. Their specialized feathers help with temperature regulation, their large wings are used for balance, and their powerful kicks serve as a potent defense mechanism.
Summary
Details
Additional Information
The flightless ostrich is the world's largest bird. They roam African savanna and desert lands and get most of their water from the plants they eat.
Speed and Movement
Though they cannot fly, ostriches are fleet, strong runners. They can sprint up to 43 miles an hour and run over distance at 31 miles an hour. They may use their wings as "rudders" to help them change direction while running. An ostrich's powerful, long legs can cover 10 to 16 feet in a single stride. These legs can also be formidable weapons. Ostrich kicks can kill a human or a potential predator like a lion. Each two-toed foot has a long, sharp claw.
Herds and Reproduction
Ostriches live in small herds that typically contain less than a dozen birds. Alpha males maintain these herds, and mate with the group's dominant hen. The male sometimes mates with others in the group, and wandering males may also mate with lesser hens. All of the group's hens place their eggs in the dominant hen's nest—though her own are given the prominent center place. The dominant hen and male take turns incubating the giant eggs, each one of which weighs as much as two dozen chicken eggs.
Behavior and Diet
Contrary to popular belief, ostriches do not bury their heads in the sand. The old saw probably originates with one of the bird's defensive behaviors. At the approach of trouble, ostriches will lie low and press their long necks to the ground in an attempt to become less visible. Their plumage blends well with sandy soil and, from a distance, gives the appearance that they have buried their heads in the sand.
Ostriches typically eat plants, roots, and seeds but will also eat insects, lizards, or other creatures available in their sometimes harsh habitat.
Details:
Scientific classification
Kingdom : Animalia
Phylum : Chordata
Class : Aves
Infraclass : Palaeognathae
Order : Struthioniformes
Family : Struthionidae
Genus : Struthio
Ostriches are large flightless birds. Two living species are recognised; the common ostrich, native to large parts of Sub-Saharan Africa, and the Somali ostrich, native to the Horn of Africa.
They are the heaviest and largest living birds, with adult common ostriches weighing anywhere between 63.5 and 145 kilograms and laying the largest eggs of any living land animal. With the ability to run at 70 km/h (43.5 mph), they are the fastest birds on land. They are farmed worldwide, with significant industries in the Philippines and in Namibia. South Africa produces about 70% of global ostrich products, with the industry largely centered around the town of Oudtshoorn. Ostrich leather is a lucrative commodity, and the large feathers are used as plumes for the decoration of ceremonial headgear. Ostrich eggs and meat have been used by humans for millennia. Ostrich oil is another product that is made using ostrich fat.
Ostriches are of the genus Struthio in the order Struthioniformes, part of the infra-class Palaeognathae, a diverse group of flightless birds also known as ratites that includes the emus, rheas, cassowaries, kiwi and the extinct elephant birds and moa.
The common ostrich was historically native to the Arabian Peninsula, and ostriches were present across Asia as far east as China and Mongolia during the Late Pleistocene and possibly into the Holocene.
Taxonomic history
The genus Struthio was first described by Carl Linnaeus in 1758. The genus was used by Linnaeus and other early taxonomists to include the emu, rhea, and cassowary, until they each were placed in their own genera. The Somali ostrich (Struthio molybdophanes) has recently become recognized as a separate species by most authorities, while others are still reviewing the evidence.
Evolution
Struthionidae is a member of the Struthioniformes, a group of paleognath birds which first appeared during the Early Eocene, and includes a variety of flightless forms which were present across the Northern Hemisphere (Europe, Asia and North America) during the Eocene epoch. The closest relatives of Struthionidae within the Struthioniformes are the Ergilornithidae, known from the late Eocene to early Pliocene of Asia. It is therefore most likely that Struthionidae originated in Asia.
The earliest fossils of the genus Struthio are from the early Miocene ~21 million years ago of Namibia in Africa, so it is proposed that genus is of African origin. By the middle to late Miocene (5–13 mya) they had spread to and become widespread across Eurasia. While the relationship of the African fossil species is comparatively straightforward, many Asian species of ostrich have been described from fragmentary remains, and their interrelationships and how they relate to the African ostriches are confusing. In India, Mongolia and China, ostriches are known to have become extinct only around, or even after, the end of the last ice age; images of ostriches have been found prehistoric Chinese pottery and petroglyphs.
Distribution and habitat
Today, ostriches are only found natively in the wild in Africa, where they occur in a range of open arid and semi-arid habitats such as savannas and the Sahel, both north and south of the equatorial forest zone. The Somali ostrich occurs in the Horn of Africa, having evolved isolated from the common ostrich by the geographic barrier of the East African Rift. In some areas, the common ostrich's Masai subspecies occurs alongside the Somali ostrich, but they are kept from interbreeding by behavioral and ecological differences. The Arabian ostriches in Asia Minor and Arabia were hunted to extinction by the middle of the 20th century, and in Israel attempts to introduce North African ostriches to fill their ecological role have failed. Escaped common ostriches in Australia have established feral populations.
Species
In 2008, S. linxiaensis was transferred to the genus Orientornis. Three additional species, S. pannonicus, S. dmanisensis, and S. transcaucasicus, were transferred to the genus Pachystruthio in 2019. Several additional fossil forms are ichnotaxa (that is, classified according to the organism's trace fossils such as footprints rather than its body) and their association with those described from distinctive bones is contentious and in need of revision pending more good material.
Additional Information
Ostrich, (Struthio camelus), is a large flightless bird found only in open country in Africa. The largest living bird, an adult male may be 2.75 metres (about 9 feet) tall—almost half of its height is neck—and weigh more than 150 kg (330 pounds); the female is somewhat smaller. The ostrich’s egg, averaging about 150 mm (6 inches) in length by 125 mm (5 inches) in diameter and about 1.35 kg (3 pounds), is also the world’s largest. The male is mostly black but has white plumes in the wings and tail; females are mostly brown. The head and most of the neck, reddish to bluish in colour, is lightly downed; the legs, including the powerful thighs, are bare. The head is small, the bill short and rather wide; the big brown eyes have thick black lashes.
Ostriches are seen individually, in pairs, in small flocks, or in large aggregations, depending on the season. The ostrich relies on its strong legs—uniquely two-toed, with the main toe developed almost as a hoof—to escape its enemies, chiefly humans and the larger carnivores. A frightened ostrich can achieve a speed of 72.5 km (45 miles) per hour. If cornered, it can deliver dangerous kicks.
Ostriches live mainly on vegetation but also take some animal food, mainly insects; they can go without water for long periods. Breeding males emit lionlike roars and hisses as they fight for a harem of three to five hens. A communal nest scraped in the ground contains more than a dozen shiny, whitish eggs. The major hen of the harem may get rid of some of the eggs to make incubation more manageable. The male sits on the eggs by night; the females take turns during the day. The chicks hatch in about 40 days and when a month old can keep up with running adults. To escape detection, chicks as well as adults may lie on the ground with neck outstretched, a habit that may have given rise to the mistaken belief that the ostrich buries its head in the sand when danger threatens. Ostrich plumes adorned the helmets of medieval European knights, and in the 19th century such plumes were sold for women’s finery. This demand led to the establishment of ostrich farms in South Africa, the southern United States, Australia, and elsewhere, but the trade collapsed after World War I. Ostriches are now raised for their meat and hide, which provides a soft, fine-grained leather. The birds have been trained for saddle and sulky racing, but they tire easily and are not well suited for training. They do well in captivity and may live 50 years.
The ostrich is typical of a group of flightless birds called ratites. Ostrich populations differing slightly in skin colour, size, and egg features formerly were considered separate species, but now they are considered to be merely races of Struthio camelus. Most familiar is the North African ostrich, S. camelus camelus, ranging, in much-reduced numbers, from Morocco to Sudan. Ostriches also live in eastern and southern Africa. The Syrian ostrich (S. camelus syriacus) of Syria and Arabia became extinct in 1941. The ostrich is the only living species in the genus Struthio. Ostriches are the only members of the family Struthionidae in the order Struthioniformes—a group that also contains kiwis, emus, cassowaries, and rheas. The oldest fossil relatives of ostriches belong to the species Calciavis grandei, which were excavated from the Green River Formation in Wyoming and date to the Eocene Epoch, some 56 million to 34 million years ago.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2584 2025-09-08 20:23:09
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2384) Seahorse
Gist
The seahorse is a marine fish of the genus Hippocampus and family Syngnathidae, named for its horse-like head and upright posture, resembling a miniature horse in the sea. These bony fish have distinctive features such as a prehensile tail, independently moving eyes, and segmented bony armour, and they are found in shallow coastal waters worldwide.
Summary
A seahorse, (genus Hippocampus), any of about 50 species of marine fishes allied to pipefishes in the family Syngnathidae (order Gasterosteiformes). Seahorses are found in shallow coastal waters in latitudes from about 52° N to 45° S. Their habitats include coral reefs, mangroves, sea grass beds, and estuaries. They are unique in appearance, with their horselike head, prehensile tail, independently moving eyes, and brood pouch. They have long, tubular snouts and small, toothless mouths. Their bodies are covered with consecutive rings of bony plates. The name of the genus that contains seahorses is taken from the Greek words hippos (meaning “horse”) and kampos (meaning “sea monster”).
Seahorses vary in size, ranging in length from about 2 to 35 cm (about 0.8 to 14 inches). Adults of some of the smallest species—such as Denise’s pygmy seahorse (Hippocampus denise), found in the tropical western Pacific from Indonesia to Vanuatu, and Satomi’s pygmy seahorse (H. satomiae), found in the tropical Indian and Pacific Oceans from the Bay of Bengal to the Coral Sea—are less than 2 cm long. The largest species, the big-bellied seahorse (H. abdominalis), which inhabits the waters off South Australia and New Zealand, can grow up to 35 cm (13.8 inches) in length.
Seahorses are rather immobile, swimming more slowly than other fishes. When swimming they maintain a vertical position and propel themselves forward using a soft-rayed dorsal fin. They use pectoral fins located on the side of the head to maneuver. Some scientists contend that this upright swimming posture evolved shortly after the expansion of sea grasses in the western Pacific roughly 25 million years ago. These plants provided seahorses with useful hiding places to avoid enemies and to capture unsuspecting prey, and ancestors of the seahorse evolved to maximize the opportunities offered by this new habitat.
Seahorses are usually found clinging to plants or corals with their tails. Their sedentary habits coupled with excellent camouflage abilities render them successful ambush predators. When small organisms swim nearby, a seahorse may capture them by rapidly sucking them into the mouth. Seahorses also rely upon camouflage to avoid predators such as crabs and other fishes.
The reproductive behaviour of seahorses is notable in that the male carries the fertilized eggs. After an elaborate courtship, the female uses an ovipositor (egg duct) to place her eggs into a brood pouch located at the base of the male’s tail where the eggs are later fertilized. Depending on the species, the eggs remain in the pouch between 10 days and six weeks. During this time the male nurtures the developing young by regulating the chemistry of the fluid inside the pouch, slowly transforming it from that of his internal body fluids to that of salt water as pregnancy progresses. To nourish the growing young, the male also produces inorganic compounds and releases the hormone prolactin, which helps break down the proteins contributed by the female. Once the eggs hatch, the male convulses his body and expels the young through a single opening in the pouch. The young are miniature versions of their parents that receive no further care. The male can receive another brood of eggs almost immediately after giving birth. In some species a male and female will maintain a monogamous pair bond throughout the breeding season and produce many broods.
Commercially, seahorses are traded live as aquarium animals and dead for use in traditional medicines and as curios. Threatened by direct overfishing, accidental capture (bycatch) in other fisheries, and the destruction of their coastal habitats, some species—such as the Pacific seahorse (H. ingens)—face extinction.
Details:
Scientific classification
Kingdom: Animalia
Phylum: Chordata
Class: Actinopterygii
Order: Syngnathiformes
Family: Syngnathidae
Subfamily: Hippocampinae
Genus: Hippocampus
A seahorse (also written sea-horse and sea horse) is any of 46 species of small marine bony fish in the genus Hippocampus. The genus name comes from the Ancient Greek hippókampos (ἱππόκαμπος), itself from híppos (ἵππος) meaning "horse" and kámpos meaning "sea monster" or "sea animal". Having a head and neck suggestive of a horse, seahorses also feature segmented bony armour, an upright posture and a curled prehensile tail. Along with the pipefishes and seadragons (Phycodurus and Phyllopteryx) they form the family Syngnathidae.
Evolution and fossil record
Anatomical evidence, supported by molecular, physical, and genetic evidence, demonstrates that seahorses are highly modified pipefish. The fossil record of seahorses, however, is very sparse. The best known and best studied fossils are specimens of Hippocampus guttulatus (though literature more commonly refers to them under the synonym of H. ramulosus), from the Marecchia River formation of Rimini Province, Italy, dating back to the Lower Pliocene, about 3 million years ago. The earliest known seahorse fossils are of two pipefish-like species, H. sarmaticus and H. slovenicus, from the coprolitic horizon of Tunjice Hills, a middle Miocene lagerstätte in Slovenia dating back about 13 million years.
Molecular dating implies that pipefish and seahorses diverged during the Late Oligocene. This has led to speculation that seahorses evolved in response to large areas of shallow water, newly created as the result of tectonic events. The shallow water would have allowed the expansion of seagrass habitats that served as camouflage for the seahorses' upright posture. These tectonic changes occurred in the western Pacific Ocean, pointing to an origin there, with molecular data suggesting two later, separate invasions of the Atlantic Ocean. In 2016, a study published in Nature found the seahorse genome to be the most rapidly evolving fish genome studied so far.
The evolution of seahorses from pipefish may have been an adaptation related to the biomechanics of prey capture. The unique posture of the seahorse allows them to capture small shrimps at larger distances than the pipefish is capable of.
Description
Seahorses range in size from 1.5 to 35 cm (0.6 to 13.8 in). They are named for their equine appearance, with bent necks and long snouted heads and a distinctive trunk and tail. Although they are bony fish, they do not have scales, but rather thin skin stretched over a series of bony plates, which are arranged in rings throughout their bodies. Each species has a distinct number of rings. The armor of bony plates also protects them against predators, and because of this outer skeleton, they no longer have ribs. Seahorses swim upright, propelling themselves using the dorsal fin, another characteristic not shared by their close pipefish relatives, which swim horizontally. Razorfish are the only other fish that swim vertically. The pectoral fins, located on either side of the head behind their eyes, are used for steering. They lack the caudal fin typical of fishes. Their prehensile tail is composed of square-like rings. They are adept at camouflage, and can grow and reabsorb spiny appendages depending on their habitat.
Unusual among fish, a seahorse has a flexible, well-defined neck. It also sports a crown-like spine or horn on its head, termed a "coronet", which is distinct for each species.
Seahorses swim extremely poorly, rapidly fluttering a dorsal fin and using pectoral fins to steer. The slowest-moving fish in the world is H. zosterae (the dwarf seahorse), with a top speed of about 1.5 m (5 ft) per hour. Since they are poor swimmers, they are most likely to be found resting with their prehensile tail wound around a stationary object. They have long snouts, which they use to drag up food, and their eyes can move independently of each other like those of a chameleon.
Habitat
Seahorses are mainly found in shallow tropical and temperate salt water throughout the world, from about 45°S to 45°N. They live in sheltered areas such as seagrass beds, estuaries, coral reefs, and mangroves. Four species are found in Pacific waters from North America to South America. In the Atlantic, Hippocampus erectus ranges from Nova Scotia to Uruguay. H. zosterae, known as the dwarf seahorse, is found in the Bahamas.
Colonies have been found in European waters such as the Thames Estuary.
Two species live in the Mediterranean Sea: H. guttulatus (the long-snouted seahorse), and H. hippocampus (the short-snouted seahorse). These species form territories; males stay within 1 sq m (10 sq ft) of habitat, while females range over about one hundred times that.
Feeding habits
Seahorses rely on stealth to ambush small prey such as copepods. They use pivot feeding to catch the copepod, which involves rotating their snout at high speed and then sucking in the copepod.
Seahorses use their long snouts to eat their food with ease. However, they are slow to consume their food and have extremely simple digestive systems that lack a stomach, so they must eat constantly to stay alive.[28] Seahorses are not very good swimmers, and for this reason they need to anchor themselves to seaweed, coral or anything else that will keep the seahorse in place. They do this by using their prehensile tails to grasp their object of choice. Seahorses feed on small crustaceans floating in the water or crawling on the bottom. With excellent camouflage, seahorses ambush prey that floats within striking range, sitting and waiting until an optimal moment. Mysid shrimp and other small crustaceans are favorites, but some seahorses have been observed eating other kinds of invertebrates and even larval fish. In a study of seahorses, the distinctive head morphology was found to give them a hydrodynamic advantage that creates minimal interference while approaching an evasive prey. Thus the seahorse can get very close to the copepods on which it preys. After successfully closing in on the prey without alerting it, the seahorse gives an upward thrust and rapidly rotates the head aided by large tendons that store and release elastic energy, to bring its long snout close to the prey. This step is crucial for prey capture, as oral suction only works at a close range. This two-phase prey capture mechanism is termed pivot-feeding. Seahorses have three distinctive feeding phases: preparatory, expansive, and recovery. During the preparatory phase, the seahorse slowly approaches the prey while in an upright position, after which it slowly flexes its head ventrally. In the expansive phase, the seahorse captures its prey by simultaneously elevating its head, expanding the buccal cavity, and sucking in the prey item. During the recovery phase, the jaws, head, and hyoid apparatus of the seahorse return to their original positions.
The amount of available cover influences the seahorse's feeding behaviour. For example, in wild areas with small amounts of vegetation, seahorses will sit and wait, but an environment with extensive vegetation will prompt the seahorse to inspect its environment, feeding while swimming rather than sitting and waiting. Conversely, in an aquarium setting with little vegetation, the seahorse will fully inspect its environment and makes no attempt to sit and wait.
Additional Information
Seahorses are members of the pipefish family. In addition to their iconic appearance, seahorses possess many interesting attributes. Among them are specialized structures in their skin cells, called chromatophores, which allow the mostly sessile seahorses to change color to mimic their surroundings. Well camouflaged as they cling to stalks of seagrass in their shallow habitats, seahorses can be hard to see.
Their truly remarkable biological claim to fame, however, is that male seahorses and sea dragons get pregnant and bear young—a unique adaptation in the animal kingdom.
After completing an elaborate courtship dance that may go on for hours or days, the female seahorse transfers her mature eggs into the male’s brood pouch, where they are fertilized. At the end of a gestation period usually lasting from two to four weeks, the pregnant male’s abdominal area begins to undulate rhythmically, and strong muscular contractions eject from a few dozen to as many as 1,000 fully formed baby seahorses into the surrounding water. After that, the offspring must fend for themselves. Large litters are necessary because only about 0.5 percent will survive to adulthood.
Many, if not all, of the 47 known seahorse species—14 of which were identified only in the 21st century—are in decline worldwide.
Because seahorses generally live in shallow, near-coastal waters, human activities including development, pollution, fisheries, and traditional medicine have reduced their numbers. At the same time, their universal appeal has worked against them; until recently, wild seahorses were often captured for the aquarium trade. The delicate creatures tend to fare poorly in aquaria, however. In recent years, captive-bred seahorses have shown promise as hardier tank-dwellers than their wild relatives.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2585 2025-09-11 21:03:40
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2385) 49th Parallel
Gist
The 49th parallel most commonly refers to the 49th parallel north, a line of latitude that forms the international border between the United States and Canada for a significant portion of their boundary, a result of treaties in 1818 and 1846. This parallel also serves as the southern border for several Canadian provinces and the northern border for various U.S. states.
Summary
The 49th parallel north is a circle of latitude that is 49° north of Earth's equator. It crosses Europe, Asia, the Pacific Ocean, North America, and the Atlantic Ocean.
The city of Paris is about 15 kilometres (9 miles) south of the 49th parallel and is the largest city between the 48th and 49th parallels. Its main airport, Charles de Gaulle Airport, lies on the 49th parallel.
Roughly 2,030 kilometres (1,260 miles) of the Canada–United States border was designated to follow the 49th parallel from British Columbia to Manitoba on the Canada side, and from Washington to Minnesota on the U.S. side, more specifically from the Strait of Georgia to the Lake of the Woods. This international border was specified in the Anglo-American Convention of 1818 and the Oregon Treaty of 1846, though survey markers placed in the 19th century cause the border to deviate from the 49th parallel by up to 810 metres (0.503 miles).
From a point on the ground at this latitude, the sun is above the horizon for 16 hours, 12 minutes during the summer solstice and 8 hours, 14 minutes during the winter solstice.
This latitude also roughly corresponds to the minimum latitude in which astronomical twilight can last all night near the summer solstice. All-night astronomical twilight lasts about from June 9 to July 2. At midnight on the summer solstice, the altitude of the sun is about −17.56°.
Slightly less than one-eighth of the Earth's surface is north of the 49th parallel.
Details
When there’s a continent at stake, arguments over borders can make or break the peace across the whole region.
What happens when a continent has only two major nations? In the case of Canada and the United States of America, the 5,525 mile long line delineating the two nations is currently the longest undefended international boundary in the world, often cited as an example to the rest of the world of how two nations can cooperate.
Today, the countries are seen as model neighbours, but it took centuries of arguing, skirmishes, and outright wars to settle the matter of who got what in North America. Even after a century and a half of relative peace, today there are still a few areas that are still in dispute, and it doesn't show signs of being settled any time soon.
If you're interested in how this comparatively simple division of land occurred, here’s how it went down:
1700s
1775 - During the Revolutionary War, the American colonists hoped Quebec and Nova Scotia would be their allies against the British. The soon-to-be U.S. got impatient, though, and took matters into its own hands, invading its neighbor to the north and taking Montreal in the first big military maneuver of the war. A month later, they were pushed back in a major defeat known as the Battle of Quebec.
1783 - After the war, a victorious U.S. demanded that Great Britain give them Canada. Unsurprisingly, the British refused. The parties settled the boundaries, or so they thought, by using the 45th parallel as the northern border between New York and New Brunswick, and creating an imaginary line through the Great Lakes. No one knew much about what existed beyond the Mississippi River to the west, so those boundaries were left murky, to say the least.
1800s
1803 - The young United States made a very good deal with France, known as the Louisiana Purchase. For 15 million dollars, they bought a vast swath of territory that reached from Gulf of Mexico to the Rockies (or Stony Mountains, as they were called then). The British and the U.S. used the watershed between the Hudson Bay and the Mississippi/Missouri rivers to establish the northern border for the newly purchased lands. That turned out to be a bad idea, because the watershed was too flat to be measured accurately.
1812 - During the War of 1812, the United States invaded Canada again, not once, but twice, as a way of attacking British interests. Some Americans thought taking Canada would be a walk in the park, but they suffered a humiliating defeat in the Siege of Detroit, aided by the Native American Tecumseh. As well, British forces land near Washington D.C., burning the US Capitol and the Whitehouse.
In 1814, a treaty restored the original boundaries. Many Canadians fought during the war, which created a new sense of a national identity.
1818 - American settlers had streamed further west, encouraged by a belief in Manifest Destiny, which held that America was meant to stretch from coast to coast. That pressured Britain and the US to return to the border-negotiating table.
Surveyors were struggling against great odds to map exactly what existed in the largely rugged terrain. The powers-that-were decided to use a straight line - the 49th parallel - to demarcate the border up to the Rockies. Left unresolved was the mysterious land beyond, called Columbia in Canada and Oregon in the U.S.
For now, Britain and the U.S allowed it to remain open to whoever could survive out there. Surveying mistakes led to an anomaly that still exists, the Northwest Angle. It’s part of Minnesota, but if you want to go there by land, you have to travel through Canada, twice.
1846 - The agreement to keep the Oregon and Columbia territories neutral was falling apart. Thousands of Americans had stakes in the region, and the U.S. was pushing hard against Britain’s Hudson Bay Company, which controlled Canadian interests. President James K. Polk demanded that U.S. territory be extended northward, past the 49th parallel.
The slogan “54° 40’ or Fight” became a rallying cry for his supporters. In the end, the U.S. blinked, and Polk agreed to let the 49th parallel become the official dividing line for the westernmost areas of the two countries, resolving (almost) the last major piece of the Canadian-U.S. puzzle. Spats over small areas would continue for decades.
1867 - Canada is granted its independence from Great Britain. Based upon a drive for self-determination from the Canadian provinces, and a desire from Britain for Canada to defend itself against American encroachment, this process was peaceful and did not lead to a military dispute as seen in America's war of Independence.
The first Canadian prime minister Sir John A. MacDonald drives the creation of a transcontinental railroad in order to allow quick movements of troops to the western territories of Canada in case of American expansion.
1900s
1908 - A treaty between the two nations is signed, establishing a joint commission tasked with surveying and delineating the border between the two neighbours.
1925 - Canada and America agree to make the International Boundary Commission permanent, in order to maintain the land and monuments along the border.
One of its responsibilities is maintenance of the Peace Arch, which was built on the exact line between the two nations, in Washington State in the U.S. and British Columbia in Canada. On the U.S side, the monument reads "Children Of a Common Mother," and on the Canadian side "Brethren Dwelling together in Unity." And for the most part, both are true.
Current
The U.S. and Canada have agreed to disagree over a few remaining bits and bobs, including Machias Seal Island in the east, which has a Canadian lighthouse but is claimed by the U.S.
There is also the matter of the Northwest Passage, which Canada says is hers, but the U.S. says is international shipping waters. A small piece off territorial waters off the coast of the Yukon (a Canadian Territory) is claimed by the U.S. as a special economic zone.
As far as we know, there are no more plans to invade, however. It was a long road, but in the end, Canada and the United States reached a harmonious relationship that stretches from the Atlantic to the Pacific.
Additional Information
The 49th parallel serves as the border between the United States and Canada due to a series of historical agreements and treaties that shaped its establishment. The boundary is drawn along the 49th parallel from Lake of the Woods in the east to the Strait of Georgia in the west. Several key events led to this decision, making it a straightforward and agreed-upon solution for both nations.
The choice of the 49th parallel as a border was both practical and symbolic. It provided a simple, horizontal line that was straightforward to draw and maintain.
This choice allowed me and others involved to easily demarcate the land and limit disputes. The decision also meant a fair division of territory between the nations, which allowed for westward expansion without constant conflict.
Despite its simplicity, drawing a border along the 49th parallel wasn’t easy. The land includes plains, mountains, and everything in between, complicating the task. Surveyors, including those on my team, faced many obstacles, like harsh weather, as they worked over several decades to accurately map and mark this boundary.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2586 2025-09-13 00:04:57
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2386) Bat (Mammal)
Gist
Mammals are a group of vertebrate animals. Examples of mammals include rats, cats, dogs, deer, monkeys, apes, bats, whales, dolphins, and humans.
Bats are mammals, and are the only mammals capable of true, sustained flight. As mammals, they are warm-blooded, covered in fur, and nourish their young with milk. Their wings are unique extensions of their forelimbs, consisting of long, spread-out fingers with a thin membrane of skin stretching between them. Bats belong to the order Chiroptera, and there are over 1,300 species worldwide.
A bat is a type of mammal from the order Chiroptera, characterized by its ability to fly using wings formed by a membrane stretched between its long fingers and body. Like other mammals, bats are warm-blooded, have fur, and nourish their young with milk. They are the only mammals capable of true, sustained flight and play vital roles in ecosystems by controlling insect pests, pollinating plants, and spreading seeds.
Summary
A bat, (order Chiroptera) is any member of the only group of mammals capable of flight. This ability, coupled with the ability to navigate at night by using a system of acoustic orientation (echolocation), has made the bats a highly diverse and populous order. More than 1,200 species are currently recognized, and many are enormously abundant. Observers have concluded, for example, that some 100 million female Mexican free-tailed bats (Tadarida brasiliensis mexicana) form summer nursery colonies in Texas, where they produce about 100 million young in five large caves. The adult males are equal in number to the females, though they do not all range as far north as Texas. Furthermore, this species is found throughout tropical America. Thus, one species alone numbers, at the very least, in the hundreds of millions.
General features
All bats have a generally similar appearance in flight, dominated by the expanse of the wings, but they vary considerably in size. The order is usually divided into two well-defined suborders: the Megachiroptera (the large Old World fruit bats) and the Microchiroptera (small bats found worldwide). Among members of the Megachiroptera, flying foxes (Pteropus) have a wingspan of 1.5 metres (about 5 feet) and a weight of 1 kg (2.2 pounds). The largest insectivorous bat is probably the naked, or hairless, bat (Cheiromeles torquatus); it weighs about 250 grams (about 9 ounces). The largest of the carnivorous bats (and the largest bat in the New World) is the spectral bat (Vampyrum spectrum), also known as the tropical American false vampire bat, with a wingspan of over 60 cm (24 inches). The tiny hog-nosed, or bumblebee, bat (Craseonycteris thonglongyai) of Thailand is one of the smallest mammals. It has a wingspan of barely 15 cm (6 inches) and weighs about 2 grams (about 0.07 ounce).
Bats vary in colour and in fur texture. Facial appearance, dominated by the muzzle and ears, varies strikingly between families and often between genera. In several families a complex fleshy adornment called the nose leaf surrounds the nostrils. Although the exact function of these facial appurtenances has yet to be determined, scientists believe they may help to direct outgoing echolocation calls (see below Orientation). Wing proportions are modified according to mode of flight. The tail and the membrane between the legs also differ, perhaps as adaptations to feeding, flight, and roosting habits. Finally, bats vary in the postures they assume when roosting, particularly in whether they hang suspended or cling to a wall and in the manner in which the wings are folded and used.
Distribution
Bats are particularly abundant in the tropics. In West Africa, for example, more than 30 genera embracing nearly 100 species have been cataloged; in the United States 19 genera, totaling about 45 species, are known. Of the 18 bat families, 3—the vesper bats (family Vespertilionidae), free-tailed bats (family Molossidae), and horseshoe bats (family Rhinolophidae)—are well represented in the temperate zones. A few American leaf-nosed bats (family Phyllostomidae) range into mild temperate regions. Several vesper bats range well into Canada.
The Vespertilionidae are found worldwide except in the polar regions and on isolated islands. The brown bats of genus Myotis have a range almost equal to that of the entire order. The free-tailed bats and sheath-tailed bats (family Emballonuridae) also encircle the Earth but are restricted to the tropics and subtropics. The horseshoe bats extend throughout the Old World, the roundleaf bats (family Hipposideridae) and Old World fruit bats (family Pteropodidae) throughout the Old World tropics, and the leaf-nosed bats throughout the New World tropics and slightly beyond. The other families have more restricted ranges.
Details
For centuries, bats have been called sinister and spooky, likely because of their beady eyes and razor-sharp fangs. But there’s more to these nocturnal creatures than meets the eyes. There are more than 1,300 species of bats in the world, making them the second most common group of mammals after rodents. Some weigh less than a penny, while others have a wingspan of six feet, but all are impressive and vital members of their ecosystems.
Winging it
The scientific name for bats is Chiroptera, which is Greek for “hand wing.” That’s because bats have four long fingers and a thumb, each connected to the next by a thin layer of skin. They are the only mammals in the world that can fly, and they are remarkably good at it. Their flexible skin membrane and movable joints allow them to change direction quickly and catch mosquitoes in midair.
Classification
There are two main types of bats: microbats and megabats. Most bats are microbats, which eat insects like moths, that come out at night. Vampire bats are the only species of microbats that feed on blood rather than insects. But not to worry—they prefer to drink from cattle and horses, not humans.
To navigate dark caves and hunt after dark, microbats rely on echolocation, a system that allows them to locate objects using sound waves. They echolocate by making a high-pitched sound that travels until it hits an object and bounces back to them. This echo tells them an object’s size and how far away it is.
In contrast, megabats live in the tropics and eat fruit, nectar, and pollen. They have larger eyes and a stronger sense of smell than microbats but have smaller ears because they don’t echolocate. There are more than 150 species of megabats, which are usually, but not always, larger than microbats.
Roosting
Bats can be found nearly everywhere, except in polar regions, extreme deserts, and a few isolated islands. They spend their daylight hours hiding in roosts around the tropics, dense forests, and wetlands. Roosts are where bats go to rest, usually in cracks and crevices that keep them hidden and protected. The most common roosts are existing structures such as caves, tree hollows, and old buildings.
Seasons often dictate where any bats choose their homes. depending on the time of year because they hibernate during the winter. For example, in the winter, some may hibernate in caves, and in the summer, they’ll return to an attic. Because good roosts can be hard to find, many live in giant colonies with millions of other bats.
No matter where they spend their seasons, all bats roost upside down. They can hang from their hind feet and legs while resting. Scientists still aren’t sure why bats do this, but here’s one theory: Bats have to fall into flight, which makes hanging upside down the best way to escape quickly.
Nature’s conservationists
Despite all the misconceptions surrounding bats, they are very important to humans and the environment. Insect-eating microbats consume millions of bugs a night, acting as a natural pest control for plants. Thanks to bats, farmers might rely less on toxic pesticides, which costs them millions of dollars each year. Nectar-drinking bats pollinate plants so they can produce fruit. In fact, more than 500 plant species, including mangoes, bananas, and avocados, depend on bats for pollination. Finally, fruit-eating bats help disperse seeds so rainforests can grow, helping to mitigate the effects of widespread deforestation.
Additional Information
Bats are flying mammals of the order Chiroptera. With their forelimbs adapted as wings, they are the only mammals capable of true and sustained flight. Bats are more agile in flight than most birds, flying with their very long spread-out digits covered with a thin membrane or patagium. The smallest bat, and arguably the smallest extant mammal, is Kitti's hog-nosed bat, which is 29–34 mm (1.1–1.3 in) in length, 150 mm (5.9 in) across the wings and 2–2.6 g (0.071–0.092 oz) in mass. The largest bats are the flying foxes, with the giant golden-crowned flying fox (Acerodon jubatus) reaching a weight of 1.6 kg (3.5 lb) and having a wingspan of 1.7 m (5 ft 7 in).
The second largest order of mammals after rodents, bats comprise about 20% of all classified mammal species worldwide, with over 1,400 species. These were traditionally divided into two suborders: the largely fruit-eating megabats, and the echolocating microbats. But more recent evidence has supported dividing the order into Yinpterochiroptera and Yangochiroptera, with megabats as members of the former along with several species of microbats. Many bats are insectivores, and most of the rest are frugivores (fruit-eaters) or nectarivores (nectar-eaters). A few species feed on animals other than insects; for example, the vampire bats are haematophagous (feeding on blood). Most bats are nocturnal, and many roost in caves or other refuges; it is uncertain whether bats have these behaviours to escape predators. Bats are distributed globally in all except the coldest regions. They are important in their ecosystems for pollinating flowers and dispersing seeds; many tropical plants depend entirely on bats for these services. Globally, they transfer organic matter into cave ecosystems and arthropod suppression. Insectivory by bats in farmland constitutes an ecosystem service that has paramount value to humans: even in today’s pesticide era, natural enemies account for almost all pest suppression in farmed ecosystems.
Bats provide humans with some direct benefits, at the cost of some disadvantages. Bat dung has been mined as guano from caves and used as fertiliser. Bats consume insect pests, reducing the need for pesticides and other insect management measures. Some bats are also predators of mosquitoes, suppressing the transmission of mosquito-borne diseases. Bats are sometimes numerous enough and close enough to human settlements to serve as tourist attractions, and they are used as food across Asia and the Pacific Rim. However, fruit bats are frequently considered pests by fruit growers. Due to their physiology, bats are one type of animal that acts as a natural reservoir of many pathogens, such as rabies; and since they are highly mobile, social, and long-lived, they can readily spread disease among themselves. If humans interact with bats, these traits become potentially dangerous to humans.
Depending on the culture, bats may be symbolically associated with positive traits, such as protection from certain diseases or risks, rebirth, or long life, but in the West, bats are popularly associated with darkness, malevolence, witchcraft, vampires, and death.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2587 2025-09-14 22:29:02
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2387) Blue Whale
Gist
Blue whales are the largest animals ever to live on our planet. They feed almost exclusively on krill, straining huge volumes of ocean water through their baleen plates (which hang from the roof of the mouth and work like a sieve). Some of the biggest individuals may eat up to 6 tons of krill a day.
There are an estimated 10,000 to 25,000 blue whales left in the world, with recent research indicating that the population may be increasing. The species is still classified as endangered by the IUCN Red List of Threatened Species due to historical whaling, but current threats include entanglement in fishing gear and vessel strikes.
Summary
The blue whale (Balaenoptera musculus) is a marine mammal and a baleen whale. Reaching a maximum confirmed length of 29.9–30.5 m (98–100 ft) and weighing up to 190–200 t (190–200 long tons; 210–220 short tons), it is the largest animal known to have ever existed. The blue whale's long and slender body can be of various shades of greyish-blue on its upper surface and somewhat lighter underneath. Four subspecies are recognized: B. m. musculus in the North Atlantic and North Pacific, B. m. intermedia in the Southern Ocean, B. m. brevicauda (the pygmy blue whale) in the Indian Ocean and South Pacific Ocean, and B. m. indica in the Northern Indian Ocean. There is a population in the waters off Chile that may constitute a fifth subspecies.
In general, blue whale populations migrate between their summer feeding areas near the poles and their winter breeding grounds near the tropics. There is also evidence of year-round residencies, and partial or age- and sex-based migration. Blue whales are filter feeders; their diet consists almost exclusively of krill. They are generally solitary or gather in small groups, and have no well-defined social structure other than mother–calf bonds. Blue whales vocalize, with a fundamental frequency ranging from 8 to 25 Hz; their vocalizations may vary by region, season, behavior, and time of day. Orcas are their only natural predators.
The blue whale was abundant in nearly all the Earth's oceans until the end of the 19th century. It was hunted almost to the point of extinction by whalers until the International Whaling Commission banned all blue whale hunting in 1966. The International Union for Conservation of Nature has listed blue whales as Endangered as of 2018. Blue whales continue to face numerous man-made threats such as ship strikes, pollution, ocean noise, and climate change.
Details
A blue whale (Balaenoptera musculus) is the most massive animal ever to have lived, a species of baleen whale that weighs approximately 150 tons and may attain a length of more than 30 metres (98 feet). The largest accurately measured blue whale was a 29.5-metre female that weighed 180 metric tons (nearly 200 short [U.S.] tons), but there are reports of 33-metre catches that may have reached 200 metric tons. Their hearts, which typically weigh about 180 kg (roughly 400 pounds), are the largest of any animal.
The blue whale is a cetacean and is classified scientifically within the order Cetacea as a rorqual (family Balaenopteridae) related to the gray whale (family Eschrichtiidae) and the right whales (Balaenidae and Neobalaenidae) of the baleen whale suborder, Mysticeti.
Blue whales are blue-gray in colour with lighter gray mottling in the form of large spots, which appear as if they were dabbed on with a huge paintbrush. The lower surfaces of the flippers are lighter gray or white in some instances. The blue whale has been called the sulfur-bottom whale because of the yellowish underside of some individuals that is reminiscent of the pale yellow colour of that chemical element; this coloration is imparted by certain algae (diatoms) living on the whale’s body. The blue whale has a wide head, a small dorsal fin located near the fluke, and 80–100 long grooves running lengthwise down the throat and chest. Its mouth contains up to 800 plates of short, wide, black baleen, or “whalebone,” with thick, coarse bristles used for catching food. Females are generally larger than males, and the largest animals live in the Southern Ocean around Antarctica.
The blue whale is found alone or in small groups in all oceans, but populations in the Southern Hemisphere are much larger. In the Northern Hemisphere, blue whales can be seen regularly in the Gulf of St. Lawrence and off the coasts of Monterey, California, and Baja California, Mexico. They spend the summer in polar waters, feeding on shrimplike crustaceans called krill. During a dive, the blue whale may engage in a series of turns and 360° rolls to locate prey and rapidly reorient its body to sweep up large concentrations of krill in a single open-mouthed lunge. A single adult blue may consume as much as eight tons of krill per day. In the winter blue whales move toward the Equator to breed. After a gestation of about 12 months, one calf about 8 metres (about 26 feet) long is born in temperate waters. While nursing, calves gain up to 90 kg (about 198 pounds) per day on the rich milk of their mothers. Young are weaned after seven to eight months, when they have reached a length of about 15 metres (about 49 feet).
Once the most important of the commercially hunted baleen whales, the blue whale was greatly reduced in numbers during the first half of the 20th century. In the 1930–31 season alone the worldwide kill of blue whales exceeded 29,000. The species has been protected from commercial whaling since the mid-1960s. Populations of blue whales appear to be recovering and are estimated worldwide at between 10,000 and 25,000 animals. However, the International Union for Conservation of Nature still lists the blue whale as an endangered species.
Additional Information
The blue whale is the largest animal on the planet, weighing as much as 200 tons (approximately 33 elephants). The blue whale has a heart the size of a Volkswagen Beetle. Its stomach can hold one ton of krill and it needs to eat about four tons of krill each day. They are the loudest animals on Earth and are even louder than a jet engine. Their calls reach 188 decibels, while a jet reaches 140 decibels. Their low frequency whistle can be heard for hundreds of miles and is probably used to attract other blue whales.
Whales are at the top of the food chain and have an important role in the overall health of the marine environment. During the 20th century, the blue whale was an important whaling target and even after it was protected and commercial whaling stopped in 1966, exploitation efforts by the former Soviet Union persisted.
Like other large whales, blue whales are threatened by environmental change including habitat loss and toxics. Blue whales can also be harmed by ship strikes and by becoming entangled in fishing gear. Although commercial whaling no longer represents a threat, climate change and its impact on krill (shrimp-like crustaceans), blue whales' major prey, makes this cetacean particularly vulnerable.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2588 2025-09-15 18:11:50
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2388) Gulf
Gulf
"Gulf" refers to a large area of sea that is partially enclosed by land, such as the Gulf of Mexico, or a wide, deep chasm or separation between things, like the "gulf between rich and poor". The term can also refer to the famous Gulf Oil company, known for its lubricants and lubricants for vehicles.
A gulf is a portion of the ocean that penetrates land. Gulfs vary greatly in size, shape, and depth. They are generally larger and more deeply indented than bays. Like bays, they often make excellent harbors.
Summary
A gulf is a large inlet from an ocean or their seas into a landmass, larger and typically (though not always) with a narrower opening than a bay. The term was used traditionally for large, highly indented navigable bodies of salt water that are enclosed by the coastline. Many gulfs are major shipping areas, such as the Persian Gulf, Gulf of Mexico, Gulf of Finland, and Gulf of Aden.
Geographical Meaning
A gulf is a coastal indentation or a portion of the ocean or sea that penetrates the land.
It is typically larger and more deeply indented than a bay.
Examples include the Persian Gulf, the Gulf of Aden, and the Gulf of Finland.
Figurative Meaning
The term "gulf" can describe a wide or impassable separation, or a significant difference between two things or groups.
For instance, there is a widening gulf between different social classes.
Details
A gulf is a portion of the ocean that penetrates land. Gulfs vary greatly in size, shape, and depth. They are generally larger and more deeply indented than bays. Like bays, they often make excellent harbors. Many important trading centers are located on gulfs.
Gulfs may be formed by movements in Earth's crust. The planet's tectonic plates may rift, or break apart, creating a gulf. Or one plate may fold under another, a process called subduction. Subduction may create a gulf by making downfolds, or troughs, in the rock under the ocean.
Gulfs are sometimes connected to the ocean by narrow passages of water called straits. Gulfs can also have wide openings and are sometimes indistinguishable from larger bodies of water.
Major Gulfs
The Gulf of Mexico, bordered by the United States, Mexico, and the island nation of Cuba, is the world's largest gulf. It has a coastline of about 5,000 kilometers (3,100 miles). The Gulf of Mexico is connected to the Atlantic Ocean by the Straits of Florida, between Cuba and the U.S. state of Florida. It is connected to the Caribbean Sea by the Yucatán Channel, between Cuba and the Mexican peninsula of Yucatán.
The Gulf of Mexico is an important economic site for all three countries. The process of upwelling occurs near the Florida coast of the gulf, creating a rich variety of sea life. Upwelling is the process in which cold, nutrient-rich water from the bottom of the gulf is brought to the surface.
Fish and other organisms thrive in areas of upwelling. Commercial, sport, and recreational fishing thrive in the Gulf of Mexico. Oil deposits sit beneath the western Gulf of Mexico. Both Mexico (in the Bay of Campeche) and the U.S. (mainly around the coasts of Texas and Louisiana) have oil wells in the Gulf of Mexico.
The Gulf Stream, one of the most powerful ocean currents in the world, originates in the Gulf of Mexico. Gulf ports, including Houston, Texas; New Orleans, Louisiana; Veracruz, Mexico; and Havana, Cuba, continue to be important cities where goods are imported and exported by sea.
The Gulf of Mexico is also the site of strong storms. Hurricanes and other storms need warm water to develop. The Gulf of Mexico is a very warm body of water, so storms can often increase their strength. Cuba and Florida are regularly hit by hurricanes on their Atlantic and Gulf coasts.
Pollution also threatens life in the Gulf of Mexico. Oil shipping and drilling can spill tons of petroleum into the ecosystem. Two huge rivers, the Mississippi in the U.S. and the Grijalva in Mexico, empty into the gulf. Chemicals used for agriculture and industry have seeped into the water, helping to create one of the largest dead zones in the world. (A dead zone is a region where there is little oxygen or life beneath the surface of the ocean.)
River management has redirected the flow of the Mississippi River. Canals, dams, and drainage systems for agriculture and industry have provided power and irrigated land. They have also reduced the wetlands at the rivers mouth and delta. The Gulfs wetlands slow storms as they move toward land. The loss of these wetlands may have contributed to the destruction brought by Hurricane Katrina to the Gulf Coast between central Florida and Texas in 2005.
The Gulf of Carpentaria, on Australia's northeast coast, is an inlet of the Arafura Sea. Because the sea and the gulf are shallow, the exchange of water between the two is reduced. Sediment collects at the mouth of the gulf, forming underwater barriers. The low shore is bordered in some areas by wetlands and swamps.
This shallow gulf with a wide mouth creates the conditions for a yearly spectacle called the Morning Glory Cloud. In September and October, sea breezes from the Gulf of Carpentaria and the Arafura Sea meet and create an enormous, fast-moving cloud over the gulf. The Morning Glory Cloud can be 1,000 kilometers (621 miles) long and move at a rate of 60 kilometers per hour (37 miles per hour).
The Persian Gulf is an arm of the Arabian Sea bordered by Iran, Iraq, Kuwait, Saudi Arabia, Qatar, Bahrain, the United Arab Emirates, and Oman. Vast deposits of petroleum in this region make the Persian Gulf strategically important. Middle Eastern countries depend on the gulf for trade and for access to the Indian Ocean. All countries that consume oil from the region, including the U.S., have a vital interest in keeping the gulf open to shipping.
Additional Information
A gulf is any large coastal indentation. More specifically, such a feature is the reentrant of an ocean, regardless of size, depth, configuration, and geologic structure. The nomenclature for gulfs is far from uniform; names that may refer to sizable gulfs in various places include bay, bight, firth, sound, and fjord. In addition, a number of pronounced concavities of oceanic margins have no proper name at all. As such, many of the characteristics of gulfs may also apply to bays and other similar geographies.
This problem of nomenclature extends to the difference between gulfs and seas. There are many small seas, such as the Sea of Marmara (11,000 square km [about 4,200 square miles]) and the Sea of Azov (38,000 square km [about 14,700 square miles]), which, strictly speaking, are really gulfs of the ocean or other seas (the Sea of Azov is a gulf of the Black Sea). The Gulf of Aden (about 270,000 square km [about 104,000 square miles]), another example, is part of the Arabian Sea, and these water bodies have a common regime (similar tides, precipitation, evaporation, and so forth). The narrow sound of Bab el-Mandeb connects the gulf with the vast Red Sea (438,100 square km [about 169,000 square miles]) and exhibits a number of specific geomorphic features. The Red Sea in turn has two small gulfs to the north—namely, those of Suez and Aqaba.
The Bay of Bengal and the Arabian Sea are both gulfs, approximately the same size and having the same monsoonal water circulation. The Bay of Bengal is, however, the largest of the gulfs, with a surface area of 2,172,000 square km (838,600 square miles), a length of 1,850 km (1,150 miles), and a width of about 1,600 km (1,000 miles).
In some cases, the width of a gulf may exceed its length. The Great Australian Bight has the widest mouth (2,800 km [1,740 miles]). The Gulf of Guinea is the deepest; its maximum depth (6,363 metres [20,876 feet]) exceeds that of the Bay of Bengal by more than 1,000 metres (about 3,300 feet).
Topographic characteristics
Single gulfs usually are formed along linear shores of the continents. If the shoreline is irregular and has a complex geologic structure, groups of gulfs of a similar nature may occur. Most shorelines have small reentrants of various size that are called bays.
The shape and bottom topography of gulfs are amazingly diverse. They are determined by the geologic structure and development of the region. Homogeneous bedrock of low strength or resistance results in simple shapes and shallow depths. The Gulf of Riga (of the Baltic Sea) is a possible example of the type. Long, narrow arms with approximately parallel shores of the south Kara Sea extend inland for about 800 km (about 500 miles). They occupy troughs that originated by erosion during a period of lower sea level (Baidaratskaya Bay, Obskaya Bay with Tazovskaya Bay tributary, Yenisey Bay, Gydanskaya Bay). Deep, angular gulfs, on the other hand, are created along fractures, faults, and rifts (e.g., Varanger Fjord); they usually have irregular bottom topography. Parallel fractures form extremely deep, narrow gulfs with parallel shores, such as the Gulf of California. Genuine fjord-gulfs are notable for their very high length-to-width ratios (up to 50:1). In regions that have undergone nonuniform deformation and uplift, gulfs of complicated and irregular shape and bottom topography are consequently formed; the Gulf of St. Lawrence is an example.
Gulfs are connected with the sea by means of one or more straits. Sometimes there may be an archipelago in the mouth of the gulf, as in the Gulf of Bothnia. There are some gulfs that open into the sea or into another gulf on opposite sides (Baffin Bay, the Gulf of Aden, and the Gulf of Oman).
Factors that affect the characteristics of gulfs
Gulfs may differ from the adjacent ocean (or sea) by virtue of water properties and dynamics and processes of sedimentation. Such differences are determined by the size and the shape of a given gulf, by the depth and bottom topography, and, to a considerable extent, by the degree of isolation from the ocean. Climatic conditions also are important. Isolation from an adjacent ocean depends on the ratio of width of mouth to total surface area of a gulf or on the cross section of the mouth to total water volume. If there is a sill (a submarine ridge or rise), the ratio of depth above the sill to the depth of the gulf is of great importance. No extensive comparisons of these ratios have been made to date; hence, any analysis of controlling variables must remain somewhat qualitative.
A high sill hampers the water exchange between an ocean and gulf and may lead to stagnation (oxygen deficiency), as is found in some fjords of Norway, in the Red Sea, and, particularly, in the Black Sea. Also, the presence of a sill causes independent circulation of gulf waters, generated by local winds and the runoff of rivers. Sills are not indispensable for the formation of an independent circulation, however. A narrow mouth, as in the Gulf of Bothnia, leads to the same result.
In humid climates, the waters of gulfs are freshened by river runoff. Salinity is particularly low in the gulfs of the Baltic Sea and along the southern coast of the Kara Sea. Water becomes almost fresh in their heads, especially in the spring when snow begins to thaw. Gulfs of the arid zone suffer from intensive evaporation and receive little river runoff. Thus, salinity increases markedly in this climatic regime—up to 60 parts per thousand in the Persian Gulf and up to 350 parts per thousand in the Kara-Bogaz-Gol (a gulf of the Caspian Sea). In addition to its effect on salinity, river runoff delivers organic matter and nutrient salts that may determine the specific features of life in the gulfs. The number of genera and species of organisms is small, but the organisms present tend to develop in quantities. That is why shrimp, oyster, and other fisheries are concentrated in many gulfs.
Funnel-shaped gulfs, in which the depth gradually decreases headward, usually have resonant tides. The tidal range at the head of such gulfs is several times greater than that in the open ocean (e.g., Bristol Channel, Río de la Plata, Mezenskaya Bay, Shelikhova Gulf). The world maximum tidal range has been registered in the Bay of Fundy (18 metres [59 feet]). The regularity (magnitude and frequency) of the flood tide may be distorted in such instances, and the duration of the flood tide may become much shorter than that of the ebb tide. This may cause the phenomenon of tidal bore, in which a steep wave will move rapidly upstream for dozens of kilometres.
Gulfs of simple shape with a narrow mouth and a high degree of isolation from the ocean are often subject to seiches. These free oscillations can result from rapid changes of atmospheric pressure and, of course, from tectonic movements such as earthquakes. Seiches gradually decrease, but some oscillation continues long after their cause disappears. A high rise of the water (storm surge) occurs in long and shallow gulfs if winds from the sea are protracted. Such phenomena are difficult to predict, and the high water levels may cause floods. Seiches commonly occur at the heads of Helgoländer Bay in the North Sea and in the Gulf of Finland.
Certain aspects of sedimentation are affected by the isolation of gulfs from the ocean and river runoff. The rate of sediment accumulation in gulfs of limited area may be very high. This, of course, is a function of river discharge; sediment composition is usually similar to that of the load transported by entering rivers. Deposition of calcium carbonate often occurs in shallow gulfs in the arid zones where few if any perennial streams exist. The bottoms of long gulfs (or gulfs having sills) are usually covered with silt even at the shallowest depths (e.g., Hudson Bay, the Bo Hai [Gulf of Chihli], the inlets or gubas of the Kara Sea, the Gulf of Riga). Only strong tidal currents can prevent this siltation and, in some cases, cause the opposite phenomenon of bottom erosion. Currents maintain the existence of or actively deepen bottom troughs in narrow-mouthed gulfs whose depths are more than 200 metres (about 660 feet), whereas depths of adjacent parts of the open ocean are only on the order of some dozens of metres.
Waves of the open ocean either do not penetrate into comparatively isolated gulfs or—if they do—they become greatly reduced after entry. Small local waves that are related to gulf size prevail there. This tends to make gulfs quite navigable, and seaports and harbours have generally been situated on them.
Classification of gulfs
The geologic structure and developmental history of gulfs are as varied as are those of the continents or oceans proper. The factors discussed above influence the morphological peculiarities of gulfs, and the latter in turn permit some general division or classification of these features to be made. The several groups in one possible scheme are discussed here using typical gulfs of each group as examples.
Areas situated in open concavities of the continental coast (Gulf of Alaska, Bay of Biscay, Gulf of Guinea, Great Australian Bight, Bay of Bengal, Gulf of Tehuantepec, for example) are classified as the A1 group. The depth of these gulfs in the region of the mouth usually is on the order of kilometres. The continental shelf and continental slope are generally pronounced. The general shape of such gulfs is simple; width of mouth usually exceeds its length. Water circulation and its physical properties are similar to those of the oceans. The character of the marine faunas does not differ from that of oceanic areas.
Large areas considerably isolated from oceans, such as the Gulf of Mexico and Baffin Bay, are designated as group A2. The former includes a geosynclinal hollow, founded in the Mesozoic Era (251 million to 65.5 million years ago) and finally shaped during the Paleogene and Neogene periods (65.5 million to 2.6 million years ago). It is connected with the ocean by the narrow and relatively shallow Straits of Florida and the Yucatán Channel. Baffin Bay is a rift hollow that is connected by straits with the Atlantic.
Ocean gulfs, such as the Gulfs of Oman, California, Aden, and some others, have smaller areas and are isolated to a lesser degree. These features, in group A3, have shapes that are determined by young faults and fractures. Depths in these gulfs generally exceed 1 kilometre (0.6 mile). Unlike the previous group, in which gulfs might be of composite geologic structure, these occupy areas that have undergone only a single episode of deformation.
Gulfs situated on the continental shelf, such as the Bay of Fundy, Hudson Bay, Río de la Plata, San Matías Gulf (off Argentina), and others, are in group B. The depth of such gulfs is up to 200 metres (about 660 feet) or more, and their configuration is determined by geologic conditions. Because shelf areas repeatedly became dry land when the sea level fell during the ice ages, these gulfs received their final shape during the Pleistocene Epoch. The Gulf of St. Lawrence is included in this group, though it is really intermediate between groups A3 and B. It contains both a pronounced shelf and a long trough up to 530 metres (1,740 feet) deep.
Gulfs of intercontinental and marginal seas are considered to be a third category. These may be divided into group C1, which consists of gulfs of basin seas, including the deepwater part only (Gulf of Aqaba) or both the deepwater and the shelf parts (Gulf of Honduras), and group C2, the shelf gulfs of the same seas (e.g., the Persian Gulf, the Gulf of Suez, Anadyrsky Gulf, the Bristol and Norton channels, and Shelikhova Gulf).
Finally, there are the gulfs of the shelf seas (gubas of the Arctic seas of Russia, gulfs of the Baltic and the White Seas, the Gulf of Carpentaria, the Bo Hai, and many others), which are placed in group D. The shallow character of the shelf seas influences the water dynamics of the gulfs. Water exchange is weakened, and sediments may accumulate in the gulf mouths, thus forming submarine barriers and further reducing exchange.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2589 2025-09-16 16:38:30
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2389) Vatican City
Gist
Vatican City is the world's smallest sovereign state, an independent city-state and the administrative center of the Roman Catholic Church, located as an enclave within Rome, Italy. Ruled by the Pope as the head of the Holy See, it is a site of immense religious and cultural importance, home to landmarks like St. Peter's Basilica, the Vatican Museums, and the Sistine Chapel. Established by the Lateran Treaty of 1929, it serves as the global spiritual headquarters for the Catholic Church and is a pilgrimage site for Christians worldwide.
Vatican City is famous for being the sovereign spiritual and administrative center of the Roman Catholic Church, the world's smallest independent state, and a major pilgrimage site. It is home to significant religious and cultural landmarks like St. Peter's Basilica and the Sistine Chapel with Michelangelo's frescoes, as well as the extensive Vatican Museums with its vast collection of art, all of which draw millions of tourists and pilgrims annually.
Summary
Vatican City, officially the Vatican City State, often shortened as the Vatican, is a landlocked sovereign state and city-state. Ruled by the pope, it is an enclave within Rome and serves as the administrative centre of the Catholic Church. Vatican City is governed by the See of Rome, commonly known as the Holy See, itself a sovereign entity under international law, which maintains its temporal power, governance, diplomacy, and spiritual independence. Vatican is also used as a metonym for the Holy See, which is the central governing body of the Catholic Church and Vatican City, comprising the pope and the Roman Curia. The independent state of Vatican City came into existence in 1929 via the Lateran Treaty between the Holy See and the Kingdom of Italy, which spoke of it as a new creation, not as a vestige of the much larger Papal States (756–1870), which had previously encompassed much of Central Italy.
With an area of 49 hectares (121 acres)[g] and a population of about 882 in 2024, it is the smallest sovereign state in the world both by area and by population. It is among the least populated capitals in the world. As governed by the Holy See, Vatican City State is an ecclesiastical or sacerdotal-monarchical state ruled by the pope, who is the bishop of Rome and head of the Catholic Church; the highest state functionaries are all Catholic clergy of various origins. The Holy See dates to early Christianity and is the principal episcopal see of the Catholic Church, which in 2018 had about 1.329 billion baptized Catholics in the world, in the Latin Church and 23 Eastern Catholic Churches. After the Avignon Papacy (1309–1377) the popes have mainly resided at the Apostolic Palace within what is now Vatican City, although at times residing instead in the Quirinal Palace in Rome or elsewhere.
Vatican City contains religious and cultural sites such as St Peter's Basilica, the Sistine Chapel, the Vatican Apostolic Library, and the Vatican Museums. They feature some of the world's most famous paintings and sculptures. The economy of Vatican City is supported financially by donations from Catholic believers, by the sale of postage stamps and souvenirs, fees for admission to museums, and sales of publications. Vatican City has no taxes, and items are duty-free.
Details
Vatican City is a landlocked ecclesiastical state, seat of the Roman Catholic Church, and an enclave surrounded by Rome, situated on the west bank of the Tiber River. Vatican City is the world’s smallest fully independent nation-state.
Layout of the city
Vatican City’s medieval and Renaissance walls form its boundaries, except on the southeast at St. Peter’s Square (Piazza San Pietro). Of the six entrances, only three—the piazza, the Arco delle Campane (Arch of the Bells) in the facade of St. Peter’s Basilica, and the entrance to the Vatican Museums and Galleries in the north wall—are open to the public. The most imposing building is St. Peter’s Basilica, built during the 4th century and rebuilt during the 16th century. Erected over the tomb of St. Peter the Apostle, it is the second largest religious building (after Yamoussoukro Basilica) in Christendom.
The Vatican Palace is the residence of the pope within the city walls. The Holy See is the name given to the government of the Roman Catholic Church, which is led by the pope as the bishop of Rome. As such, the Holy See’s authority extends over Catholics throughout the world. Since 1929 it has resided in Vatican City, which was established as an independent state to enable the pope to exercise his universal authority.
Vatican City has its own telephone system, post office, gardens, astronomical observatory, radio station, banking system, and pharmacy, as well as a contingent of Swiss Guards responsible for the personal safety of the pope since 1506. Almost all supplies—including food, water, electricity, and gas—must be imported. There is no income tax and no restriction on the import or export of funds. As the Holy See, it derives its income from the voluntary contributions of more than one billion Roman Catholics worldwide, as well as interest on investments and the sale of stamps, coins, and publications. Banking operations and expenditures have been reported publicly since the early 1980s.
History and governance
The city of Rome has been an important center of Christianity since the early days of the church. St. Peter, considered the first pope, is thought to have lived and died in Rome. In 313 Emperor Constantine I issued the Edict of Milan, which ended official persecution of Christians and opened the door to the growth of the church in both spiritual and material terms. By the 4th century the church had gained control of a great deal of territory, called the Patrimony of St. Peter, in and around Rome. Papal influence in central Italy began to increase in the 5th century, as the Roman Empire fell apart and the people of the area began to rely on the pope for protection from invading armies. By about the year 600 the church was one of the largest landowners in the world.
The legal basis for the foundation of the Papal States was provided by the Donation of Pippin, which granted the pope the rights to large parts of central Italy in 754. In the 9th century the first city walls (Leonine Walls) were completed under Pope Leo IV. Between the 12th and 14th centuries the Vatican underwent something of a building boom as a new palace was built and the Leonine Walls were restored. The Vatican fell into decay after 1309, when the office of the papacy was moved to Avignon in France. The pope’s return to Rome in 1377 marked the beginning of a revitalization.
Italy became a unified country in the 19th century, which led to major changes in the Vatican’s political status. Most immediately, the church lost its land to the new country. Some of the papal territories voted to join the Kingdom of Sardinia in 1859. Italy annexed the rest of the Papal States by 1870 and made Rome the Italian capital. To protest the incorporation into a unified Italy, each pope thereafter remained a voluntary “prisoner of the Vatican,” never leaving the small territory of the papal grounds. This situation lasted nearly 60 years.
In 1929 a solution to this ongoing problem was found. Vatican City’s independent sovereignty was recognized by the Fascist Italian government in the Lateran Treaty. Sovereignty is exercised by the pope upon his election as the head of the Roman Catholic Church. He has absolute executive, legislative, and judicial powers within the city. While most of the inhabitants of Vatican City are priests or nuns, they also include several hundred laypersons engaged in secretarial, domestic, trade, and service occupations.
Institutions and attractions
Special extraterritorial privileges are extended to more than 10 other buildings in Rome and to Castel Gandolfo, the pope’s summer residence in the Alban Hills. In addition, Vatican City maintains embassies in numerous foreign nations.
The Vatican enjoyed a cultural golden age during the Renaissance, when the popes were among Italy’s foremost patrons of the arts. The Vatican Museums and Galleries, the frescoes by Michelangelo in the Sistine Chapel, the frescoes by Pinturicchio in the Borgia Apartment, and Raphael’s Stanze (“Rooms”) attract critics, artists, and flocks of tourists from throughout the world. Years of restoration work on the Sistine Chapel frescoes were completed in 1994, making it possible to view Michelangelo’s work in full vibrant colors. In 2000 the millennial Jubilee focused world attention on Vatican City.
The Vatican Apostolic Library contains a priceless collection of some 150,000 manuscripts and 1.6 million printed books, many from pre-Christian and early Christian times. The Vatican publishes its own influential daily newspaper, L’Osservatore Romano, and its press can print books and pamphlets in any of 30 languages, from old Ecclesiastical Georgian to Tamil. Since 1983 the Vatican has produced its own television programming. Its radio broadcasts are heard in some 40 languages in many parts of the world. Vatican City was designated a UNESCO World Heritage site in 1984. Pop. (2019 est.) 453.
Additional Information
The Vatican City, one of the most sacred places in Christendom, attests to a great history and a formidable spiritual venture. A unique collection of artistic and architectural masterpieces lie within the boundaries of this small state. At its centre is St Peter's Basilica, with its double colonnade and a circular piazza in front and bordered by palaces and gardens. The basilica, erected over the tomb of St Peter the Apostle, is the largest religious building in the world, the fruit of the combined genius of Bramante, Raphael, Michelangelo, Bernini and Maderno.
Outstanding Universal Value:
Brief synthesis
One of the most sacred places in Christendom, Vatican City stands as a testimony to a history of about two millennia and to a formidable spiritual venture. Site of the tomb of the Apostle Saint Peter, first of the uninterrupted succession of Roman Pontiffs, and therefore a main pilgrimage centre, the Vatican is directly and tangibly linked with the history of Christianity. Furthermore, it is both an ideal and an exemplary creation of the Renaissance and of Baroque art. It exerted an enduring influence on the development of the arts from the 16th century.
The independent State, defined by the Lateran Treaty of 11 February 1929, extends its territorial sovereignty over an area of 44 ha in the centre of Rome: Vatican City enclosed by its walls and open toward the city through Bernini’s colonnade of Saint Peter’s. The boundaries of the city-state contain masterpieces and living institutions that are a witness to the unique continuity of the crucial role played by this place in the history of mankind. The Centre of Christianity since the foundation of Saint Peter’s Basilica by Constantine (4th century), and at a later stage the permanent seat of the Popes, the Vatican is at once the pre-eminently holy city for Catholics, an important archaeological site of the Roman world and one of the major cultural reference points of both Christians and non-Christians.
Its prestigious history explains the development of an architectural and artistic ensemble of exceptional value. Beneath the basilica of Saint Peter, reconstructed in the 16th century under the guidance of the most brilliant architects of the Renaissance, remains of the first basilica founded by Constantine still exist, as well as ruins of the circus of Caligula and Nero, and a Roman necropolis of the 1st century AD, where Saint Peter’s tomb is located. Under Julius II’s patronage in 1506, an extraordinary artistic era was inaugurated, leading to the decoration of Raphael’s Stanze and of the Sistine Chapel with frescoes by Michelangelo, along with the building of the new basilica, completed in 1626, fruit of the combined genius of Bramante, Raphael, Michelangelo, Bernini, Maderno and Della Porta.
The Vatican Palace is the result of a long series of additions and modifications by which, from the Middle Ages, the Popes rivalled each other in magnificence. The original building of Nicholas III (1277-1280) was enlarged in the 15th, 16th and 17th centuries: the history of the arts of the Renaissance and Baroque periods finds here iconic models.
In 1475, Sixtus IV founded the Vatican Library, which is the first open to the public in Europe; the collections of manuscripts and books, prints, drawings, coins and decorative arts, constantly increased through the centuries, making it an invaluable repository of human culture.
From the mid-18th century, the popes’ efforts were also directed towards expanding the private collections of antiquities dating back to the Renaissance: their transformation into public museums accessible to scholars and connoisseurs marks the origin of the Vatican Museums. New buildings were built specifically to house the classical sculptures, such as the Pio-Clementine Museum, which represents a milestone in the history of European culture. The 19th- and 20th-century additions of new and diverse collections and buildings accord with the tradition of papal patronage.
Criterion (i): The Vatican, a continuous artistic creation whose progress spreads over centuries, represents a unique masterpiece of the modelling of a space, integrating creations which are among the most renowned of mankind: not only the world famous icon of sacred architecture, the basilica of Saint Peter, but also the chapel of Nicholas V decorated by Fra Angelico, the Borgia apartment with frescoes by Pinturicchio, the Stanze of Raphael and his students, the Sistine Chapel, whose mural decoration, begun by Perugino, Botticelli and other painters, was completed in the 16th century with the frescoes of the ceiling and the monumental Last Judgement by Michelangelo, who left his last murals in the Pauline Chapel.
Criterion (ii): The Vatican exerted a deep influence on the development of art from the 16th century. Architects have visited it to study the constructions of Bramante (the Basilica of Saint Peter, the Belvedere Court), of Michelangelo (the cupola of Saint Peter), of Bernini (Saint Peter's colonnade, the Baldacchino of the Basilica). Both within and outside Europe, the Vatican buildings have been abundantly copied and imitated, the paintings (the frescoes of Raphael and Michelangelo) and the antiquities of the Museums no less so.
Criterion (iv): The Vatican is both an ideal and exemplary religious and palatial creation of the Renaissance and of Baroque art.
Criterion (vi): Site of the tomb of Saint Peter and pilgrimage centre, the Vatican is directly and materially linked with the history of Christianity. For more than a thousand years, mankind has accumulated, in this privileged site, the treasures of its collective memory (manuscripts and books of the Library) and of its universal genius.
Integrity
The boundaries of the property, which coincide with the entire territory of the Vatican City State, have preserved their original integrity and characteristics. The exceptional urban, architectural and aesthetic values, even through successive additions and changes in form and design, invariably maintain the highest standards of artistic quality and workmanship, building an organic ensemble of unparalleled harmony. Civil and sacred buildings, which have been in use for centuries, maintain their religious, cultural, institutional and diplomatic functions unaltered.
Authenticity
The property meets the required conditions of authenticity, since most of its features are still preserved and maintained in their initial form, perform their primary functions and truthfully convey their original spiritual and cultural values. The extensive restoration campaigns conducted on some of the most significant monuments of the site since the date of the inscription ensure the material conservation of the heritage and strengthen its capacity for expressing its values.
Protection and management requirements
The property is safeguarded by the law for the protection of the cultural heritage (no. 355, 25/07/2001) and by several rules of procedure issued by the various institutions of the Holy See in charge of heritage. For instance, the body responsible for the preservation and maintenance of Saint Peter’s Basilica, the Fabbrica di S.Pietro, was founded in 1506 and is still active. The legal protective mechanism and traditional management system are adequate and ensure the effective protection of the site. The state of conservation of the property is constantly and carefully monitored, with special attention paid to the impact of the huge number of pilgrims and visitors.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2590 2025-09-17 16:33:49
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2390) Anaconda
Gist
Is anaconda a real snake?
Yes, an anaconda is a very real, giant, non-venomous snake, but it's not a single species. Anacondas are large, heavy constrictors from tropical South America that are excellent swimmers. The largest species, the green anaconda, is the world's heaviest snake and can grow to impressive lengths.
Anacondas are semi aquatic snakes found in tropical South America, notable in the Amazon and different to pythons. They are some of the largest snakes in the world and are known for their swimming ability. “Anaconda” is the common name for the genus Eunectes, a genus of boa.
Summary
Anaconda, (genus Eunectes), any of three to five species of large, constricting, water-loving snakes found in tropical South America classified in the family Boidae (see also boa). Green anacondas (Eunectes akayima, the northern green anaconda, and E. murinus, the southern green anaconda) are among the largest snakes in the world, growing up to 9 meters (29.5 feet) long—rivaling the reticulated python (Python reticulatus) in length—and weighing up to 250 kg (550 pounds). The yellow, or southern, anaconda (E. notaeus), however, is much smaller, adult females reaching a maximum length of about 4.4–4.6 meters (roughly 14.4–15.1 feet) long. Historically, two additional forms, the beni (E. beniensis) and the dark-spotted anaconda (E. deschauenseei), which are closely related to E. notaeus, have been classified as separate species; however, growing morphological and genetic evidence suggests that both should be classified as yellow anacondas.
Natural history
Green anacondas—also called giant anacondas and common anacondas—are olive-colored snakes with alternating oval-shaped black spots. Despite much attention paid to a handful of colossal individuals, most adults do not exceed 5 meters (16 feet) in length. This group was divided formally into two species in 2024 based on the discovery of genetic differences between northern and southern populations. Northern green anacondas inhabit lowland basins, swamps, and even some drier grassy areas east of the Andes Mountains, extending from eastern Ecuador, northern Peru, and northern Brazil northward to Colombia, Venezuela, Guyana, Suriname, French Guiana, and Trinidad and Tobago. Southern green anacondas, which prefer marshes and other wet areas to drier habitats, are found from eastern Peru and northern Brazil south through northern Bolivia, extreme eastern Paraguay, and southern Brazil. Yellow anacondas inhabit swamps, marshes, and other aquatic habitats from the Paraguay River basin southward to northeastern Argentina; the species is distinguished from others by its yellowish coloration, which is broken up by pairs of overlapping black spots. Dark-spotted anacondas inhabit the seasonally flooded lower reaches of the Amazon River and coastal watersheds, extending between the Marowijne (or Maroni) River, separating Suriname and French Guiana southeast to the Tocantins River in Brazil. Benis are found primarily in seasonally flooded areas of north-central Bolivia.
Anacondas exhibit sexual dimorphism, females being larger and heavier than males. In general, they are carnivorous apex predators that consume fish and other vertebrates (including caimans and other reptiles, mammals, and waterfowl). These snakes can seize a large animal by the neck and throw their coils around it, killing it by constriction. They also kill smaller prey, such as small turtles and diving birds, with the mouth and sharp backward-pointing teeth alone. Kills made onshore are often dragged into the water, perhaps to avoid attracting jaguars, which prey on anacondas, and to ward off biting ants attracted to the carcass. Yellow anacondas target smaller vertebrate prey while also feeding on carrion, whereas green anacondas lie in the water (generally at night) to ambush larger prey, such as large caimans and mammals (including capybaras, deer, tapirs, and peccaries) that come to drink near the shoreline. Although anacondas occasionally attack people, encounters between them are rare.
Very little is known about the reproductive habits of the beni and the dark-spotted anaconda. Green and yellow anacondas, however, are polyandrous, a mating system in which single females mate with multiple males. In green and yellow anacondas, this can occur in a writhing mass of individuals, called a breeding ball, that can last several weeks. This behavior is not universal. Many females spend the breeding season, which spans the months April and May, with only one male. After mating concludes, some female green anacondas may cannibalize one or more of their male suitors (see also cannibalism, animal). Anacondas do not lay eggs. Rather, females incubate the eggs within their bodies for up to seven months before giving birth to live young. On average, female green anacondas produce about 30 live young, and female yellow anacondas give birth to about 40. The young snakes receive no parental investment, and they become sexually mature between the ages of three and four years. Anacondas live for about 10–20 years in the wild and up to 30 years in captivity.
Conservation status
Four species of anacondas are classified as species of least concern by the International Union for Conservation of Nature and Natural Resources (IUCN), and the conservation status of the northern green anaconda has yet to be assessed. The population trends of each species are not well known, but herpetologists note that these snakes are likely threatened in at least some parts of their ranges by habitat loss—due to the draining of wetland habitats for agriculture—and hunting. The yellow anaconda, the beni, and the dark-spotted anaconda are hunted for their meat and hide, and the yellow anaconda, the beni, and both species of green anaconda are targeted by people out of fear or in retaliation for attacks on livestock. Green anacondas appear to bear the brunt of this persecution because of their menacing size.
Details
Anacondas or water boas are a group of large boas of the genus Eunectes. They are a semiaquatic group of snakes found in tropical South America. Three to five extant and one extinct species are currently recognized, including one of the largest snakes in the world, E. murinus, the green anaconda.
Description
Although the name applies to a group of snakes, it is often used to refer only to one species, in particular, the common or green anaconda (Eunectes murinus), which is the largest snake in the world by weight, and the second longest after the reticulated python.
Distribution and habitat
Found in tropical South America from Ecuador, Brazil, Colombia and Venezuela south to Argentina.
Feeding
All five species are aquatic snakes that prey on other aquatic animals, including fish, river fowl, and caiman. Videos exist of anacondas preying on domestic animals such as goats and sometimes even young jaguars that venture too close to the water.
Relationship with humans
While encounters between people and anacondas may be dangerous, they do not regularly hunt humans. Nevertheless, threat from anacondas is a familiar trope in comics, movies, and adventure stories (often published in pulp magazines or adventure magazines) set in the Amazon jungle. Local communities and some European explorers have given accounts of giant anacondas, legendary snakes of much greater proportion than any confirmed specimen.
Although charismatic, there is little known on the biology of wild anacondas. Most of our knowledge comes from the work of Dr. Jesús A. Rivas and his team working in the Venezuelan Llanos.
Indigenous mythology
According to the founding myth of the Huni Kuin, a man named Yube fell in love with an anaconda woman and was turned into an anaconda as well. He began to live with her in the deep world of waters. In this world, Yube discovered a hallucinogenic drink with healing powers and access to knowledge. One day, without telling his anaconda wife, Yube decided to return to the land of men and resume his old human form. The myth also explains the origin of cipó or ayahuasca—a hallucinogenic drink taken ritualistically by the Huni Kuin.
Additional Information
Some of the largest snakes in the world, Anacondas are known for their swimming ability and there are many types.
Anacondas are semi aquatic snakes found in tropical South America, notable in the Amazon and different to pythons. They are some of the largest snakes in the world and are known for their swimming ability. “Anaconda” is the common name for the genus Eunectes, a genus of boa. Eunectes means “good swimmer” in Greek, according to SeaWorld.
There are four recognized species of anaconda, according to Bill Heyborne, a herpetologist and professor of biology at Southern Utah University. They are the green anaconda, the yellow or Paraguayan anaconda, the dark-spotted anaconda and the Beni or Bolivian anaconda. "They can be differentiated from one another genetically, but also based on their size and geographic range," Heyborne said.
Heyborne said that when most people say anaconda, they are actually referring to the green anaconda, the largest of the four species. The green anaconda is the heaviest snake in the world and one of the longest.
According to the Mythology.net , anacondas feature prominently in South American myths, sometimes appearing as shapeshifters , as the creator of the water, as vicious human-eaters, or as magical, spiritual beings with healing properties.
There have also been reports of anacondas reaching lengths of 40, 50 and 100 feet (12, 15 and 30 meters) — far longer than anything scientifically verified. These reports have given rise to the Giant Anaconda myth, popularized in the early 20th century by explorers and colonists, according to Loren Coleman and Jerome Clark's "Cryptozoology A to Z".
Anacondas are stocky, muscular snakes that are thicker than other boas, according to Reptiles Cove. They have thick necks and narrow but large heads. All anacondas have nostrils and eyes on the tops of their heads, which allow them to see above the water while remaining mostly submerged. They have a thick black stripe that runs from the eye to the jaw.
Anacondas have small, smooth scales that grow larger toward the posterior of their bodies. They have loose, soft skin that can handle a great deal of water absorption, according to the University of Michigan’s Animal Diversity Web (ADW).
Near their cloacal region, anacondas have spurs on their scales. Males have larger spurs than females, though females are overall larger and longer snakes. In fact, anacondas exhibit the largest sexual dimorphism (with the female being larger) of any tetrapod species, according to Jesus Rivas, a herpetologist and founder of the Anaconda Project.
Anacondas’ coloring and size depends on the species. Their spotted, green, yellow and brown color palettes allow them to blend in with tropical rivers and rainforests, according to Sciencing.
True to their name, these are greenish-brown, olive, or greenish-gray. They have black or brown egg-shaped spots on the mid-to posterior back of their bodies. Their sides are sometimes more yellow than green with egg-shaped spots with yellow centers, according to the Jacksonville Zoo.
Green anacondas’ length is still a matter of debate, said Heyborne. They are quite difficult to measure. It is hard to stretch out a captive anaconda, not to mention potentially dangerous for the snake, according to Rivas. People who see anacondas in the wild are likely to overestimate their length due to fear. Also, an anaconda that has recently eaten will look much larger than one that hasn’t, causing exaggerated estimations of size. Furthermore, the skins of dead snakes can be stretched, meaning that it is hard to scientifically verify the length of those, too, especially historical samples, according to Wonderpolis.
While many publications, including National Geographic and the San Diego Zoo list anacondas’ maximum verified length as 29 or 30 feet (9 m), Rivas, who has captured and measured more than 1,000 anacondas, believes they don’t grow much longer than 20 feet (6 m). The Guinness Book of World Records lists the longest snake ever recorded as a 25-foot (7.6 m) reticulated python.
The average size of female anacondas is around 15 feet (4.5 m), and the average size of males is around 9 feet (2.7 m), according to Boas and Pythons of the World.
Weights are also not known for sure. Estimates top out at about 550 lbs. (250 kilograms), according to National Geographic, but the average is probably somewhere between 100 and 150 lbs. (45 and 68 kg), according to "The Lives of Amphibians and Reptiles in an Amazonian Rainforest" (Cornell, 2005) by William E. Duellman.
Yellow anacondas have yellow, golden-tan, or yellow-green coloring with black or dark brown blotches, spots, streaks, and dorsal bands. Each snake has a unique pattern of yellow and black scales on the bottom of its tail. The average length is about 9 feet (2.7 m), according to the World Land Trust.
Not much is known about these anacondas, which were long considered a hybrid of yellow and green anacondas until scientists determined they were their own species. Their coloring is similar to the green anaconda, according to World Atlas

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2591 2025-09-18 17:19:21
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2391) Lithosphere
Gist
The lithosphere is Earth's rigid, outermost shell, encompassing the crust and the uppermost part of the mantle, broken into tectonic plates that slowly move over the asthenosphere. This rocky, brittle layer is where geological phenomena like mountain building, earthquakes, and volcanism occur, as well as where all terrestrial and marine life forms exist.
The lithosphere is the rigid, solid outer layer of the Earth, consisting of the crust and the uppermost part of the mantle. It is the layer of rock, minerals, and soil that forms the Earth's surface, and it is broken into large pieces called tectonic plates.
The lithosphere is Earth's rigid, outermost shell, encompassing the crust and the uppermost part of the mantle, broken into tectonic plates that slowly move over the asthenosphere. This rocky, brittle layer is where geological phenomena like mountain building, earthquakes, and volcanism occur, as well as where all terrestrial and marine life forms exist.
Summary
A lithosphere is the rigid, outermost rocky shell of a terrestrial planet or natural satellite. On Earth, it is composed of the crust and the lithospheric mantle, the topmost portion of the upper mantle that behaves elastically on time scales of up to thousands of years or more. The crust and upper mantle are distinguished on the basis of chemistry and mineralogy.
Earth's lithosphere
Earth's lithosphere, which constitutes the hard and rigid outer vertical layer of the Earth, includes the crust and the lithospheric mantle (or mantle lithosphere), the uppermost part of the mantle that is not convecting. The layer below the lithosphere is called the asthenosphere, which is the weaker, hotter, and deeper part of the upper mantle that is able to convect. The lithosphere–asthenosphere boundary is defined by a difference in response to stress. The lithosphere remains rigid for very long periods of geologic time in which it deforms elastically and through brittle failure, while the asthenosphere deforms viscously and accommodates strain through plastic deformation.
Due to this definition of the lithosphere– asthenosphere boundary, the thickness of the lithosphere is considered to be the depth to the isotherm associated with the transition between brittle and viscous behavior. The temperature at which olivine becomes ductile (~1,000 °C or 1,830 °F) is often used to set this isotherm because olivine is generally the weakest mineral in the upper mantle.
The lithosphere is subdivided horizontally into tectonic plates, which often include terranes accreted from other plates.
Details
The lithosphere is the solid, outer part of Earth. The lithosphere includes the brittle upper portion of the mantle and the crust, the outermost layers of Earth’s structure. It is bounded by the atmosphere above and the asthenosphere (another part of the upper mantle) below.
Although the rocks of the lithosphere are still considered elastic, they are not viscous. The asthenosphere is viscous, and the lithosphere-asthenosphere boundary (LAB) is the point where geologists and rheologists—scientists who study the flow of matter—mark the difference in ductility between the two layers of the upper mantle. Ductility measures a solid material’s ability to deform or stretch under stress. The lithosphere is far less ductile than the asthenosphere.
There are two types of lithosphere: oceanic lithosphere and continental lithosphere. Oceanic lithosphere is associated with oceanic crust, and is slightly denser than continental lithosphere.
Plate Tectonics
The most well-known feature associated with Earth’s lithosphere is tectonic activity. Tectonic activity describes the interaction of the huge slabs of lithosphere called tectonic plates.
The lithosphere is divided into tectonic plates including the North American, Caribbean, South American, Scotia, Antarctic, Eurasian, Arabian, African, Indian, Philippine, Australian, Pacific, Juan de Fuca, Cocos, and Nazca.
Most tectonic activity takes place at the boundaries of these plates, where they may collide, tear apart, or slide against each other. The movement of tectonic plates is made possible by thermal energy (heat) from the mantle part of the lithosphere. Thermal energy makes the rocks of the lithosphere more elastic.
Tectonic activity is responsible for some of Earth's most dramatic geologic events: earthquakes, volcanoes, orogeny (mountain-building), and deep ocean trenches can all be formed by tectonic activity in the lithosphere.
Tectonic activity can shape the lithosphere itself: Both oceanic and continental lithospheres are thinnest at rift valleys and ocean ridges, where tectonic plates are shifting apart from one another.
How the Lithosphere Interacts with Other Spheres
The cool, brittle lithosphere is just one of five great “spheres” that shape the environment of Earth. The other spheres are the biosphere (Earth’s living things); the cryosphere (Earth’s frozen regions, including both ice and frozen soil); the hydrosphere (Earth’s liquid water); and the atmosphere (the air surrounding our planet). These spheres interact to influence such diverse elements as ocean salinity, biodiversity, and landscape.
For instance, the pedosphere is part of the lithosphere made of soil and dirt. The pedosphere is created by the interaction of the lithosphere, atmosphere, cryosphere, hydrosphere, and biosphere. Enormous, hard rocks of the lithosphere may be ground down to powder by the powerful movement of a glacier (cyrosphere). Weathering and erosion caused by wind (atmosphere) or rain (hydrosphere) may also wear down rocks in the lithosphere. The organic components of the biosphere, including plant and animal remains, mix with these eroded rocks to create fertile soil—the pedosphere.
The lithosphere also interacts with the atmosphere, hydrosphere, and cryosphere to influence temperature differences on Earth. Tall mountains, for example, often have dramatically lower temperatures than valleys or hills. The mountain range of the lithosphere is interacting with the lower air pressure of the atmosphere and the snowy precipitation of the hydrosphere to create a cool or even icy climate zone. A region’s climate zone, in turn, influences adaptations necessary for organisms of the region’s biosphere.
Additional Information
The lithosphere is the solid shell of the planet Earth. This means the crust, plus the part of the upper mantle which behaves elastically over long time scales.
Under the lithosphere is the asthenosphere, the weaker, hotter, and deeper part of the upper mantle. This part can flow.
The lithosphere provides a conductive lid on top of the convecting mantle: it reduces heat transport through the Earth. A lithosphere (for "rocky", and for "sphere") is the rigid, outermost shell of a terrestrial-type planet or natural satellite which is defined by its rigid mechanical properties. On Earth, it is composed of the crust and that portion of the upper mantle which behaves elastically on time scales of thousands of years or greater. The outermost shell of a rocky planet, the crust, is defined on the basis of its chemistry and mineralogy.
Types of lithosphere :-
There are two types of lithosphere:-
a) Oceanic lithosphere, which is associated with oceanic crust and exists in the ocean basins. Oceanic lithosphere is typically about 50–100 km thick
b) Continental lithosphere, which is associated with continental crust. Continental lithosphere has a range in thickness from about 40 km to perhaps 200 km, of which about 40 km is crust.
The lithosphere is divided into tectonic plates, which move gradually relative to one another.
Oceanic lithosphere thickens as it ages and moves away from the mid-ocean ridge. This thickening occurs by conductive cooling, which converts hot asthenosphere into lithospheric mantle, and causes the oceanic lithosphere to become increasingly dense with age. Oceanic lithosphere is less dense than asthenosphere for a few tens of millions of years, but after this becomes increasingly dense than asthenosphere.
When a continental plate comes together with an oceanic plate, at a subduction zones, the oceanic lithosphere always sinks beneath the continental.
New oceanic lithosphere is constantly being produced at mid-ocean ridges and is recycled back to the mantle at subduction zones. As a result, oceanic lithosphere is much younger than continental lithosphere: the oldest oceanic lithosphere is about 200 million years old, while parts of the continental lithosphere are billions of years old.
Another distinguishing characteristic of the lithosphere is its flow properties. Under the influence of the low-intensity, long-term stresses that drive tectonic motion, the lithosphere is like a rigid shell. It changes mainly by breaking ("brittle failure"). The asthenosphere (the layer of the mantle below the lithosphere) is heat-softened and adjusts by plastic changes ("deformation").
Oceanic lithosphere is made up of mostly basalt and gabbro. Continental lithosphere is made up of granite and gneiss.
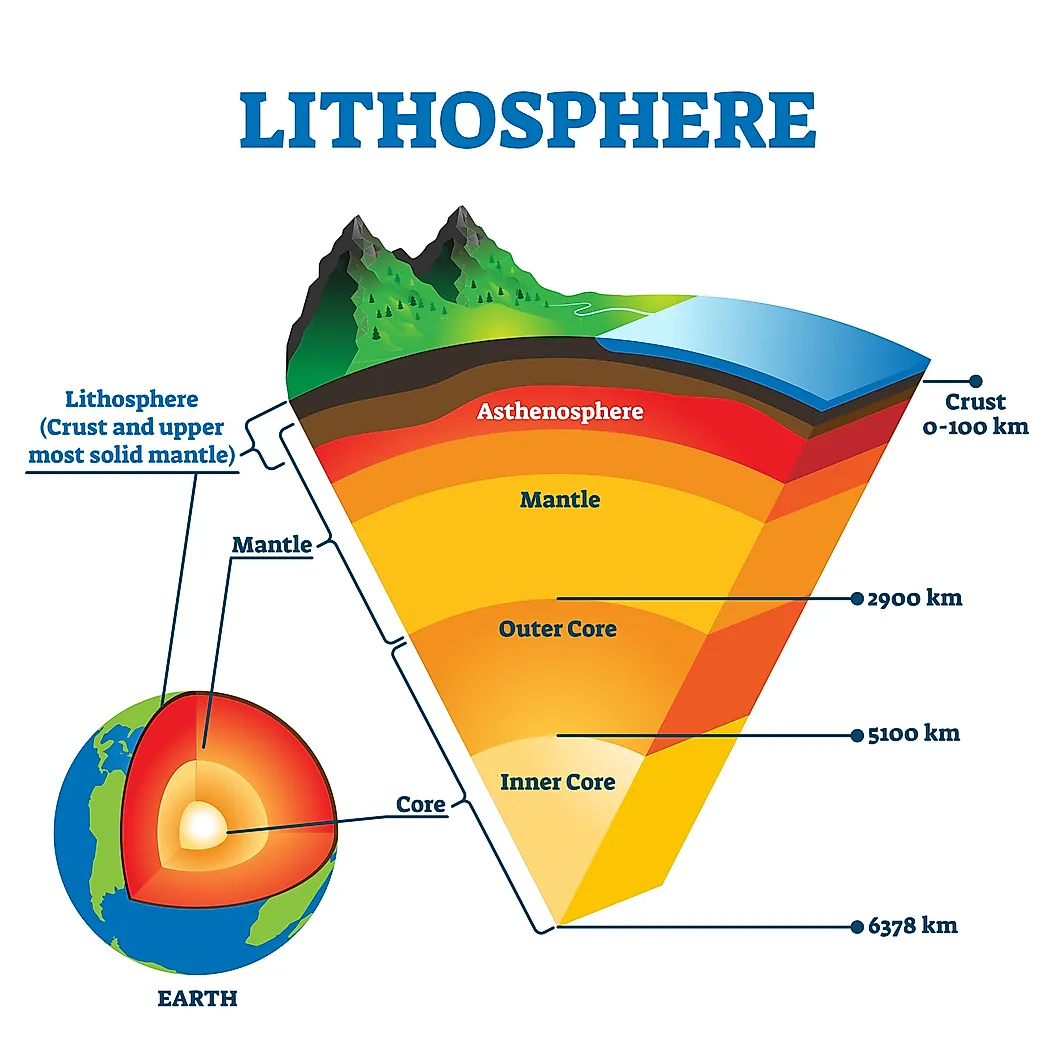
It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2592 2025-09-19 17:08:17
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2392) Hydrosphere
Gist
The hydrosphere is the sum of all the Earth's water, covering about 71% of the planet's surface. This includes oceans, rivers, lakes, groundwater, and atmospheric water vapor and ice. The hydrosphere plays a critical role in the water cycle, moving water through different states and locations, and is essential for all forms of life on Earth.
The hydrosphere is the sum of Earth's water, in the ocean, the ground, on the surface, and in the air. Approximately 71 percent of Earth's surface is covered in water. Of all of that water, only about three percent is freshwater. An even smaller amount can be used as drinking water.
Summary
The hydrosphere is the combined body of water found on, under, and above the surface of a planet, minor planet, or natural satellite. Although Earth's hydrosphere has been around for about 4 billion years, it continues to change in shape. This is caused by seafloor spreading and continental drift, which rearranges the land and ocean.
It has been estimated that there are 1.386 billion cubic kilometres (333 million cubic miles) of water on Earth. This includes water in gaseous, liquid and frozen forms as soil moisture, groundwater and permafrost in the Earth's crust (to a depth of 2 km); oceans and seas, lakes, rivers and streams, wetlands, glaciers, ice and snow cover on Earth's surface; vapour, droplets and crystals in the air; and part of living plants, animals and unicellular organisms of the biosphere. Saltwater accounts for 97.5% of this amount, whereas fresh water accounts for only 2.5%. Of this fresh water, 68.9% is in the form of ice and permanent snow cover in the Arctic, the Antarctic and mountain glaciers; 30.8% is in the form of fresh groundwater; and only 0.3% of the fresh water on Earth is in easily accessible lakes, reservoirs and river systems.
The total mass of Earth's hydrosphere is about 1.4 × {10}^{18} tonnes, which is about 0.023% of Earth's total mass. At any given time, about 2 × {10}^{13} tonnes of this is in the form of water vapor in the Earth's atmosphere (for practical purposes, 1 cubic metre of water weighs 1 tonne). Approximately 71% of Earth's surface, an area of some 361 million square kilometres (139.5 million square miles), is covered by ocean. The average salinity of Earth's oceans is about 35 grams of salt per kilogram of sea water (3.5%).
Details
Hydrosphere is a discontinuous layer of water at or near Earth’s surface. It includes all liquid and frozen surface waters, groundwater held in soil and rock, and atmospheric water vapour.
Water is the most abundant substance at the surface of Earth. About 1.4 billion cubic km (326 million cubic miles) of water in liquid and frozen form make up the oceans, lakes, streams, glaciers, and groundwaters found there. It is this enormous volume of water, in its various manifestations, that forms the discontinuous layer, enclosing much of the terrestrial surface, known as the hydrosphere.
Central to any discussion of the hydrosphere is the concept of the water cycle (or hydrologic cycle). This cycle consists of a group of reservoirs containing water, the processes by which water is transferred from one reservoir to another (or transformed from one state to another), and the rates of transfer associated with such processes. These transfer paths penetrate the entire hydrosphere, extending upward to about 15 km (9 miles) in Earth’s atmosphere and downward to depths on the order of 5 km (3 miles) in its crust.
This article examines the processes of the water cycle and discusses the way in which the various reservoirs of the hydrosphere are related through the water cycle. It also describes the biogeochemical properties of Earth’s waters at some length and considers the distribution of global water resources and their use and pollution by human society. Details concerning the major water environments that make up the hydrosphere are provided in the articles ocean, lake, river, and ice. See also climate for specific information about the impact of climatic factors on the water cycle. The principal concerns and methods of hydrology and its various allied disciplines are summarized in Earth sciences.
Distribution and quantity of Earth’s waters
Ocean waters and waters trapped in the pore spaces of sediments make up most of the present-day hydrosphere. The total mass of water in the oceans equals about 50 percent of the mass of sedimentary rocks now in existence and about 5 percent of the mass of Earth’s crust as a whole. Deep and shallow groundwaters constitute a small percentage of the total water locked in the pores of sedimentary rocks—on the order of 3 to 15 percent. The amount of water in the atmosphere at any one time is trivial, equivalent to roughly 13,000 cubic km (about 3,100 cubic miles) of liquid water, or about 0.001 percent of the total at Earth’s surface. This water, however, plays an important role in the water cycle.
At present, ice locks up a little more than 2 percent of Earth’s water and may have accounted for as much as 3 percent or more during the height of the glaciations of the Pleistocene Epoch (2.6 million to 11,700 years ago). Although water storage in rivers, lakes, and the atmosphere is small, the rate of water circulation through the rain-river-ocean-atmosphere system is relatively rapid. The amount of water discharged each year into the oceans from the land is approximately equal to the total mass of water stored at any instant in rivers and lakes.
Soil moisture accounts for only 0.005 percent of the water at Earth’s surface. It is this small amount of water, however, that exerts the most direct influence on evaporation from soils. The biosphere, though primarily H2O in composition, contains very little of the total water at the terrestrial surface, only about 0.00004 percent, yet the biosphere plays a major role in the transport of water vapour back into the atmosphere by the process of transpiration.
Biogeochemical properties of the hydrosphere:
Rainwater
About 107,000 cubic km (nearly 25,800 cubic miles) of rain fall on land each year. The total water in the atmosphere is 13,000 cubic km, and this water, owing to precipitation and evaporation, turns over every 9.6 days. Rainwater is not pure but rather contains dissolved gases and salts, fine-ground particulate material, organic substances, and even bacteria. The sources of the materials in rainwater are the oceans, soils, fertilizers, air pollution, and fossil fuel combustion.
It has been observed that rains over oceanic islands and near coasts have ratios of major dissolved constituents very close to those found in seawater. The discovery of the high salt content of rain near coastlines was somewhat surprising because sea salts are not volatile, and it might be expected that the process of evaporation of water from the sea surface would “filter” out the salts. It has been demonstrated, however, that a large percentage of the salts in rain is derived from the bursting of small bubbles at the sea surface due to the impact of rain droplets or the breaking of waves, which results in the injection of sea aerosol into the atmosphere. This sea aerosol evaporates, with resultant precipitation of the salts as tiny particles that are subsequently carried high into the atmosphere by turbulent winds. These particles may then be transported over continents to fall in rain or as dry deposition.
River and ocean waters
River discharge constitutes the main source for the oceans. Seawater has a more uniform composition than river water. It contains, by weight, about 3.5 percent dissolved salts, whereas river water has only 0.012 percent. The average density of the world’s oceans is roughly 2.75 percent greater than that of typical river water. Of the average 35 parts per thousand salts of seawater, sodium and chlorine make up almost 30 parts, and magnesium and sulfate contribute another four parts. Of the remaining one part of the salinity, calcium and potassium constitute 0.4 part each and carbon, as carbonate and bicarbonate, about 0.15 part. Thus, nine elements (hydrogen, oxygen, sulfur, chlorine, sodium, magnesium, calcium, potassium, and carbon) make up 99 percent of seawater, though most of the 94 naturally occurring elements have been detected therein. Of importance are the nutrient elements phosphorus, nitrogen, and silicon, along with such essential micronutrient trace elements as iron, cobalt, and copper. These elements strongly regulate the organic production of the world’s oceans.
In contrast to ocean water, the average salinity of the world’s rivers is low—only about 0.012 percent, or 120 ppm by weight. Of this salt content, carbon as bicarbonate constitutes 58 parts, or 48 percent, and calcium, sulfur as sulfate, and silicon as dissolved monomeric silicic acid make up a total of about 39 parts, or 33 percent. The remaining 19 percent consists predominantly of chlorine, sodium, and magnesium in descending importance. It is obvious that the concentrations and relative proportions of dissolved species in river waters contrast sharply with those of seawater. Thus, even though seawater is derived in part by the chemical differentiation and evaporation of river water, the processes involved affect every element differently, indicating that simple evaporation and concentration are entirely secondary to other processes.
Generally speaking, the composition of river water, and thus that of lakes, is controlled by water-rock interactions. The attack of carbon dioxide-charged rain and soil waters on the individual minerals in continental rocks leads to the production of dissolved constituents for lakes, rivers, and streams. It also gives rise to solid alteration products that make up soils or suspended particles in freshwater aquatic systems. The carbon dioxide content of rain and soil waters is of particular importance in weathering processes. The pH of rainwater equilibrated with the atmospheric carbon dioxide partial pressure of 10−3.5 (0.00032) atmosphere is 5.6. In industrial regions, rainwater pH values may be lower because of the release and subsequent hydrolysis of acid gases—namely, sulfur dioxide and nitrogen oxides (NOx) from the combustion of fossil fuels. After rainwater enters soils, its characteristics change markedly. The usual few parts per million of salts in rainwater increase substantially as the water reacts. The upper part of the soil is a zone of intense biochemical activity. The bacterial population near the surface is large, but it decreases rapidly downward. One of the major biochemical processes of the bacteria is the oxidation of organic material, which leads to the release of carbon dioxide. Soil gases obtained above the zone of water saturation may contain 10 to 40 times as much carbon dioxide as the free atmosphere, and in some cases carbon dioxide has been shown to make up 30 percent of the soil gases as opposed to 0.03 percent of the free atmosphere. In addition to the acid effects of carbon dioxide, there is a highly acidic microenvironment created by the roots of living plants. Values of pH as low as 2 have been measured immediately adjacent to root hairs. The combined length of a plant’s root hairs may be several kilometres, so their chemical effects on acidic water are formidable.
Additional Information
A hydrosphere is the total amount of water on a planet. The hydrosphere includes water that is on the surface of the planet, underground, and in the air.
A hydrosphere is the total amount of water on a planet. The hydrosphere includes water that is on the surface of the planet, underground, and in the air. A planet's hydrosphere can be liquid, vapor, or ice.
On Earth, liquid water exists on the surface in the form of oceans, lakes, and rivers. It also exists below ground—as groundwater, in wells and aquifers. Water vapor is most visible as clouds and fog.
The frozen part of Earth's hydrosphere is made of ice: glaciers, ice caps and icebergs. The frozen part of the hydrosphere has its own name, the cryosphere.
Water moves through the hydrosphere in a cycle. Water collects in clouds, then falls to Earth in the form of rain or snow. This water collects in rivers, lakes and oceans. Then it evaporates into the atmosphere to start the cycle all over again. This is called the water cycle.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2593 2025-09-20 18:16:42
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2393) Atmosphere
Gist
The atmosphere is a layer of gases held by gravity around a planet or other celestial body, providing air to breathe, protecting against harmful solar radiation, and regulating temperature to create a habitable environment. Earth's atmosphere consists primarily of nitrogen and oxygen, with trace amounts of other gases like argon, carbon dioxide, and water vapor. It is crucial for weather, climate, and the overall biosphere.
The atmosphere is the layer or set of layers of gases surrounding a planet, star, or other body, held in place by the body's gravity. For Earth, it is a mixture of gases, primarily nitrogen and oxygen, that provides air to breathe, shields the planet from harmful solar radiation, and helps regulate its temperature by trapping heat.
Summary
One of the main components of Earth’s interdependent physical systems is the atmosphere. An atmosphere is made of the layers of gases surrounding a planet or other celestial body. Earth’s atmosphere is composed of about 78% nitrogen, 21% oxygen, and one percent other gases. These gases are found in atmospheric layers (troposphere, stratosphere, mesosphere, thermosphere, and exosphere) defined by unique features such as temperature and pressure. The atmosphere protects life on earth by shielding it from incoming ultraviolet (UV) radiation, keeping the planet warm through insulation, and preventing extremes between day and night temperatures. The sun heats layers of the atmosphere causing it to convect driving air movement and weather patterns around the world.
Details
The atmosphere of Earth consists of a layer of mixed gas that is retained by gravity, surrounding the Earth's surface. It contains variable quantities of suspended aerosols and particulates that create weather features such as clouds and hazes. The atmosphere serves as a protective buffer between the Earth's surface and outer space. It shields the surface from most meteoroids and ultraviolet solar radiation, reduces diurnal temperature variation – the temperature extremes between day and night, and keeps it warm through heat retention via the greenhouse effect. The atmosphere redistributes heat and moisture among different regions via air currents, and provides the chemical and climate conditions that allow life to exist and evolve on Earth.
By mole fraction (i.e., by quantity of molecules), dry air contains 78.08% nitrogen, 20.95% oxygen, 0.93% argon, 0.04% carbon dioxide, and small amounts of other trace gases (see Composition below for more detail). Air also contains a variable amount of water vapor, on average around 1% at sea level, and 0.4% over the entire atmosphere.
Earth's primordial atmosphere consisted of gases accreted from the solar nebula, but the composition changed significantly over time, affected by many factors such as volcanism, outgassing, impact events, weathering and the evolution of life (particularly the photoautotrophs). In the present day, human activity has contributed to atmospheric changes, such as climate change (mainly through deforestation and fossil-fuel–related global warming), ozone depletion and acid deposition.
The atmosphere has a mass of about 5.15 × {10}^{18} kg, three quarters of which is within about 11 km (6.8 mi; 36,000 ft) of the surface. The atmosphere becomes thinner with increasing altitude, with no definite boundary between the atmosphere and outer space. The Kármán line at 100 km (62 mi) is often used as a conventional definition of the edge of space. Several layers can be distinguished in the atmosphere based on characteristics such as temperature and composition, namely the troposphere, stratosphere, mesosphere, thermosphere (formally the ionosphere), and exosphere. Air composition, temperature and atmospheric pressure vary with altitude. Air suitable for use in photosynthesis by terrestrial plants and respiration of terrestrial animals is found within the troposphere.
The study of Earth's atmosphere and its processes is called atmospheric science (aerology), and includes multiple subfields, such as climatology and atmospheric physics. Early pioneers in the field include Léon Teisserenc de Bort and Richard Assmann. The study of the historic atmosphere is called paleoclimatology.
Composition
The three major constituents of Earth's atmosphere are nitrogen, oxygen, and argon. Water vapor accounts for roughly 0.25% of the atmosphere by mass. In the lower atmosphere, the concentration of water vapor (a greenhouse gas) varies significantly from around 10 ppm by mole fraction in the coldest portions of the atmosphere to as much as 5% by mole fraction in hot, humid air masses, and concentrations of other atmospheric gases are typically quoted in terms of dry air (without water vapor). The remaining gases are often referred to as trace gases, among which are other greenhouse gases, principally carbon dioxide, methane, nitrous oxide, and ozone. Besides argon, other noble gases, neon, helium, krypton, and xenon are also present. Filtered air includes trace amounts of many other chemical compounds.
Many substances of natural origin may be present in locally and seasonally variable small amounts as aerosols in an unfiltered air sample, including dust of mineral and organic composition, pollen and spores, sea spray, and volcanic ash. Various industrial pollutants also may be present as gases or aerosols, such as chlorine (elemental or in compounds), fluorine compounds, and elemental mercury vapor. Sulfur compounds such as hydrogen sulfide and sulfur dioxide (SO2) may be derived from natural sources or from industrial air pollution.
The average molecular weight of dry air, which can be used to calculate densities or to convert between mole fraction and mass fraction, is about 28.946 or 28.964 g/mol. This is decreased when the air is humid.
Up to an altitude of around 100 km (62 mi), atmospheric turbulence mixes the component gases so that their relative concentrations remain the same. There exists a transition zone from roughly 80 to 120 km (50 to 75 mi) where this turbulent mixing gradually yields to molecular diffusion. The latter process forms the heterosphere where the relative concentration of lighter gases increase with altitude.
Stratification
In general, air pressure and density decrease with altitude in the atmosphere. However, temperature has a more complicated profile with altitude and may remain relatively constant or even increase with altitude in some regions. Because the general pattern of the temperature/altitude profile, or lapse rate, is constant and measurable by means of instrumented balloon soundings, the temperature behavior provides a useful metric to distinguish atmospheric layers. This atmospheric stratification divides the Earth's atmosphere into five main layers with these typical altitude ranges:
* Exosphere: 700–10,000 km (435–6,214 mi)
* Thermosphere: 80–700 km (50–435 mi)
* Mesosphere: 50–80 km (31–50 mi)
* Stratosphere: 12–50 km (7–31 mi)
* Troposphere: 0–12 km (0–7 mi).
Additional Information
Atmosphere is the gas and aerosol envelope that extends from the ocean, land, and ice-covered surface of a planet outward into space. The density of the atmosphere decreases outward, because the gravitational attraction of the planet, which pulls the gases and aerosols (microscopic suspended particles of dust, soot, smoke, or chemicals) inward, is greatest close to the surface. Atmospheres of some planetary bodies, such as Mercury, are almost nonexistent, as the primordial atmosphere has escaped the relatively low gravitational attraction of the planet and has been released into space. Other planets, such as Venus, Earth, Mars, and the giant outer planets of the solar system, have retained an atmosphere. In addition, Earth’s atmosphere has been able to contain water in each of its three phases (solid, liquid, and gas), which has been essential for the development of life on the planet.
The evolution of Earth’s current atmosphere is not completely understood. It is thought that the current atmosphere resulted from a gradual release of gases both from the planet’s interior and from the metabolic activities of life-forms—as opposed to the primordial atmosphere, which developed by outgassing (venting) during the original formation of the planet. Current volcanic gaseous emissions include water vapour (H2O), carbon dioxide (CO2), sulfur dioxide (SO2), hydrogen sulfide (H2S), carbon monoxide (CO), chlorine (Cl), fluorine (F), and diatomic nitrogen (N2; consisting of two atoms in a single molecule), as well as traces of other substances. Approximately 85 percent of volcanic emissions are in the form of water vapour. In contrast, carbon dioxide is about 10 percent of the effluent.
During the early evolution of the atmosphere on Earth, water must have been able to exist as a liquid, since the oceans have been present for at least three billion years. Given that solar output four billion years ago was only about 60 percent of what it is today, enhanced levels of carbon dioxide and perhaps ammonia (NH3) must have been present in order to retard the loss of infrared radiation into space. The initial life-forms that evolved in this environment must have been anaerobic (i.e., surviving in the absence of oxygen). In addition, they must have been able to resist the biologically destructive ultraviolet radiation in sunlight, which was not absorbed by a layer of ozone as it is now.
Once organisms developed the capability for photosynthesis, oxygen was produced in large quantities. The buildup of oxygen in the atmosphere also permitted the development of the ozone layer as O2 molecules were dissociated into monatomic oxygen (O; consisting of single oxygen atoms) and recombined with other O2 molecules to form triatomic ozone molecules (O3). The capability for photosynthesis arose in primitive forms of plants between two and three billion years ago. Previous to the evolution of photosynthetic organisms, oxygen was produced in limited quantities as a by-product of the decomposition of water vapour by ultraviolet radiation.
The current molecular composition of Earth’s atmosphere is diatomic nitrogen (N2), 78.08 percent; diatomic oxygen (O2), 20.95 percent; argon (A), 0.93 percent; water (H20), about 0 to 4 percent; and carbon dioxide (CO2), 0.04 percent. Inert gases such as neon (Ne), helium (He), and krypton (Kr) and other constituents such as nitrogen oxides, compounds of sulfur, and compounds of ozone are found in lesser amounts.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2594 2025-09-21 17:05:12
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2394) Arctic Circle
Gist
The Arctic Circle is an imaginary line of latitude at approximately 66.5° N, marking the southern boundary of the Arctic region. It is significant because the area north of the circle experiences periods of continuous daylight during the summer solstice (the "midnight sun") and continuous darkness during the winter solstice. This phenomenon is due to Earth's axial tilt of about 23.5 degrees.
The Arctic Circle is the northernmost region on Earth, encompassing areas north of the 66° 33' line of latitude, which includes the Arctic Ocean and the geographic North Pole.
Summary
Arctic Circle, parallel, or line of latitude around Earth, is at approximately 66°30′ N. Because of Earth’s inclination of about 23 1/2° to the vertical, it marks the southern limit of the area within which, for one day or more each year, the sun does not set (about June 21) or rise (about December 21). The length of continuous day or night increases northward from one day on the Arctic Circle to six months at the North Pole. The Antarctic Circle is the southern counterpart of the Arctic Circle, where on any given date conditions of daylight or darkness are exactly opposite.
Details
The Arctic Circle is one of the two polar circles, and the northernmost of the five major circles of latitude as shown on maps of Earth at about 66° 34' N. Its southern counterpart is the Antarctic Circle.
The Arctic Circle marks the southernmost latitude at which, on the winter solstice in the Northern Hemisphere, the Sun does not rise all day, and on the Northern Hemisphere's summer solstice, the Sun does not set. These phenomena are referred to as polar night and midnight sun respectively, and the further north one progresses, the more obvious this becomes. For example, in the Russian port city of Murmansk, three degrees north of the Arctic Circle, the Sun stays below the horizon for 20 days before and after the winter solstice, and above the horizon for 20 days before and after the summer solstice.
The position of the Arctic Circle is not fixed and currently runs 66°33′50.6″ north of the Equator. Its latitude depends on Earth's axial tilt, which fluctuates by a margin of some 2° over a 41,000-year period, due to tidal forces resulting from the orbit of the Moon. Consequently, the Arctic Circle is currently drifting northwards at a speed of about 14.5 m (48 ft) per year.
Midnight sun and polar night
The Arctic Circle is the southernmost latitude in the Northern Hemisphere at which the centre of the Sun can remain continuously above or below the horizon for twenty-four hours; as a result, at least once each year at any location within the Arctic Circle the centre of the Sun is visible at local midnight, and at least once the centre is not visible at local noon.
Directly on the Arctic Circle these events occur, in principle, exactly once per year: at the June and December solstices, respectively. However, because of atmospheric refraction and mirages, and also because the sun appears as a disk and not a point, part of the midnight sun is visible, on the night of the northern summer solstice, at a latitude of about 50 minutes of arc (′) (90 km (56 mi)) south of the Arctic Circle. Similarly, on the day of the northern winter solstice, part of the Sun may be seen up to about 50′ north of the Arctic Circle. That is true at sea level; those limits increase with elevation above sea level, although in mountainous regions there is often no direct view of the true horizon.
Human habitation
The largest communities north of the Arctic Circle are situated in Russia, Norway, and Sweden: Murmansk (population 295,374) and Norilsk (178,018) in Russia; Tromsø (75,638) in Norway, Vorkuta (58,133) in Russia, Bodø (52,357) and Harstad (24,703) in Norway; and Kiruna, Sweden (22,841). In Finland, the largest settlement in the immediate vicinity of the Arctic Circle is Rovaniemi (62,667), lying 6 km (4 mi) south of the line. Salekhard (51,186) in Russia is the only city in the world located directly on the Arctic Circle.
In contrast, the largest North American community north of the Arctic Circle, Sisimiut (Greenland), has approximately 5,600 inhabitants. In the United States, Utqiagvik, Alaska (formerly known as Barrow) is the largest settlement north of the Arctic Circle with about 5,000 inhabitants. The largest such community in Canada is Inuvik in the Northwest Territories, with 3,137 inhabitants.
Geography
The Arctic Circle is roughly 16,000 km (9,900 mi) in circumference. The area north of the Circle is about 20,000,000 sq km (7,700,000 sq mi) and covers roughly 4% of Earth's surface.
The Arctic Circle passes through the Arctic Ocean, the Scandinavian Peninsula, North Asia, Northern America, and Greenland. The land within the Arctic Circle is divided among eight countries: Norway, Sweden, Finland, Russia, the United States (Alaska), Canada (Yukon, Northwest Territories, and Nunavut), Denmark (Greenland), and Iceland (where it passes through the small offshore island of Grímsey).
Climate
The climate north of the Arctic Circle is generally cold, but the coastal areas of Norway have a generally mild climate as a result of the Gulf Stream, which makes the ports of northern Norway and northwest Russia ice-free all year long. In the interior, summers can be quite warm, while winters are extremely cold. For example, summer temperatures in Norilsk, Russia will sometimes reach as high as 30 °C (86 °F), while the winter temperatures frequently fall below −50 °C (−58 °F).
Additional Information
The Arctic Circle is one of the five major circles of latitude (imaginary lines around the Earth) that are often marked on maps of the Earth. This is the parallel of latitude that (in 2000) runs 66° 33' 39" north of the Equator. North of this line, the Sun never rises at the Winter Solstice and never sets at the Summer Solstice.
The Arctic Circle is one way to say what part of the Earth is in the Arctic. The North Pole is in the center of the Arctic Circle.
Countries which are partly within the Arctic Circle are:
* Russia
* Canada
* Denmark (Greenland)
* United States of America (Alaska)
* Norway
* Sweden
* Finland
* Iceland (less than 1 square kilometre).
The position of the Arctic Circle is not fixed. It directly depends on the Earth's axial tilt. The axial tilt fluctuates within a margin of 2° over a 40,000-year period, mostly due to tidal forces from the orbit of the Moon. The tilt is currently diminishing, so the Arctic Circle is drifting northwards at a speed of about 15 m (49 ft) per year.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2595 2025-09-22 17:18:16
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2395) Antarctic Circle
Gist
The Antarctic Circle is the southernmost of the five major circles of latitude on Earth, located at approximately 66°33' South of the equator. It marks the northern limit of the region known as the Antarctic.
The Antarctic Circle is an imaginary line of latitude located at approximately 66.5° South of the Equator, marking the boundary of the region known as the Antarctic. It defines the southernmost extent of the Earth's temperate zones and is characterized by the phenomenon of polar day and polar night, where the sun remains continuously above or below the horizon for at least 24 hours each year.
Summary
The Antarctic Circle is one of the five major circles of latitude that mark maps of the Earth. It is the parallel of latitude at 66° 33′ 39″ south of the Equator, crossing mostly the Southern Ocean.
In the Antarctic Circle, all places have twenty-four hours of daylight on the Summer Solstice in December. In June on the Winter Solstice all places have twenty-four hours of night. There is a minimum of one whole day that the sun does not set and one whole day that the sun does not rise.
The area south of this circle is named the Antarctic, and the zone to the north is the Southern Temperate Zone.
The continent of Antarctica is a land mass that is most of the area within the Antarctic Circle. The South Pole is in the center of the Antarctic Circle.
There is no permanent population of persons south of the Antarctic Circle. There are research centers from some nations on Antarctica. Teams of scientists live in the research centers for part of the year. In past centuries some whaling stations were set up on Antarctica and some of the whalers would live there for a year or more. At least three children have been born in Antarctica. It is one of the coldest places on earth.The lowest temperature ever recorded on earth was recorded in Antarctica (-129.3).
Details
The Antarctic Circle is the most southerly of the five major circles of latitude that mark maps of Earth. The region south of this circle is known as the Antarctic, and the zone immediately to the north is called the Southern Temperate Zone. South of the Antarctic Circle, the Sun is above the horizon for 24 continuous hours at least once per year (and therefore visible at solar midnight) and the centre of the Sun (ignoring refraction) is below the horizon for 24 continuous hours at least once per year (and therefore not visible at solar noon); this is also true within the Arctic Circle, the Antarctic Circle’s counterpart in the Northern Hemisphere.
The position of the Antarctic Circle is not fixed and, not taking account of the nutation, currently runs 66°33′50.6″ south of the Equator. This figure may be slightly inaccurate because it does not allow for the effects of astronomical nutation, which can be up to 10″. Its latitude depends on the Earth's axial tilt, which fluctuates within a margin of more than 2° over a 41,000-year period, due to tidal forces resulting from the orbit of the Moon. Consequently, the Antarctic Circle is currently drifting southwards at a speed of about 14.5 m (48 ft) per year.
Midnight sun and polar night
The Antarctic Circle is the northernmost latitude in the Southern Hemisphere at which the centre of the sun can remain continuously above the horizon for twenty-four hours; as a result, at least once each year at any location within the Antarctic Circle the centre of the sun is visible at local midnight, and at least once the centre of the sun is below the horizon at local noon.
Directly on the Antarctic Circle these events occur, in principle, exactly once per year: at the December and June solstices, respectively. However, because of atmospheric refraction and mirages, and because the sun appears as a disk and not a point, part of the midnight sun may be seen on the night of the southern summer solstice up to about 50 minutes (′) (90 km (56 mi)) north of the Antarctic Circle; similarly, on the day of the southern winter solstice, part of the sun may be seen up to about 50′ south of the Antarctic Circle. That is true at sea level; those limits increase with elevation above sea level, although in mountainous regions there is often no direct view of the true horizon. Mirages on the Antarctic continent tend to be even more spectacular than in Arctic regions, creating, for example, a series of apparent sunsets and sunrises while in reality the sun remains below the horizon.
Human habitation
There is no permanent human population south of the Antarctic Circle, but there are several research stations in Antarctica operated by various nations that are inhabited by teams of scientists who rotate on a seasonal basis. In previous centuries some semi-permanent whaling stations were established on the continent, and some whalers would live there for a year or more. At least eleven children have been born in Antarctica, albeit in stations north of the Antarctic Circle.
Geography
The circumference of the Antarctic Circle is roughly 16,000 kilometres (9,900 mi). The area south of the Circle is about 20,000,000 sq km (7,700,000 sq mi) and covers roughly 4% of Earth's surface. Most of the continent of Antarctica is within the Antarctic Circle.
Additional Information
Antarctic Circle is a parallel, or line of latitude around Earth, at 66°30′ S. Because Earth’s axis is inclined about 23.5° from the vertical, this parallel marks the northern limit of the area within which, for one day or more each year, at the summer and winter solstices, the sun does not set (December 21 or 22) or rise (June 21 or 22). The length of continuous day or night increases southward from one day at the Antarctic Circle to six months at the South Pole. The South Pole is located on the central ice-covered plateau of the large continental mass, the Antarctic, which almost fills the area within the Antarctic Circle. On any date, the lengths of day and night at the Antarctic Circle are the converse of those at the Arctic Circle. The Antarctic Circle, which separates the south frigid zone from the south temperate zone, was first crossed by Capt. James Cook on January 17, 1773.
It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2596 2025-09-23 17:40:17
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2396) Rain Gauge
Gist
A rain gauge is used to measure and collect liquid precipitation, providing data for water resource management, flood and drought forecasting, agricultural planning, and climate research. This information helps farmers decide on planting and harvesting, assists hydrologists and meteorologists in studying weather patterns, and allows authorities to prepare for potential flooding.
Rainfall is measured using a rain gauge, an instrument that collects rainwater in a cylindrical container for a set period. The collected water's depth is measured with a special ruler and given in millimeters or inches, indicating the amount of rain that fell. For larger areas, automated and remote-sensing technologies like weather radar are also used to estimate rainfall patterns.
A rain gauge is a scientific instrument that measures the amount of liquid precipitation over a defined area and period of time. It consists of a collector, typically a funnel, that directs rainwater into a container, allowing for the measurement of depth in inches or millimeters, often with the use of a measuring cylinder. There are non-recording and recording types, with the latter providing continuous data on rainfall intensity through moving charts or magnetic tape.
Summary
Precipitation is the process occurs when water falls from clouds to the ground. It can happen as rain, snow, sleet, or hail. The rate of precipitation varies seasonally and regionally.
A rain gauge (also known as an udometer, pluviometer, ombrometer, and hyetometer) is an instrument used by meteorologist (scientist who study weather) and hydrologists ( scientists who study water) to gather and measure the amount of liquid precipitation over a predefined area, over a period of time. It is used for determining the depth of precipitation (usually in millimeters ) that occurs over a unit area and thus measuring rainfall amount.
In Vancouver, the rainy season is winter and very little rain falls during the summer. Rain gauges are great tools to use for monitoring changes in precipitation over time. They are used by scientists to predict how much water we will have in stored in our reservoirs and to predict how much water we can use in dryer periods.
Details
A rain gauge (also known as udometer, ombrometer, pluviometer and hyetometer) is an instrument used by meteorologists and hydrologists to gather and measure the amount of liquid precipitation in a predefined area, over a set period of time. It is used to determine the depth of precipitation (usually in mm) that occurs over a unit area and measure rainfall amount.
History
People living in India and the Ancient Greeks began to record rainfall around 400 to 500 BCE. In India the readings were correlated against expected growth. In the Arthashastra, used for example in Magadha, precise standards were set as to grain production. Each state storehouse was equipped with a rain gauge to classify land for taxation purposes. Rainfall measurement was also mentioned in the Jewish text in Palestine. In 1247, the Song Chinese mathematician and inventor Qin Jiushao invented Tianchi basin rain and snow gauges to reference rain, and snowfall measurements, as well as other forms of meteorological data.
In 1441, the Cheugugi was invented during the reign of Sejong the Great of the Joseon dynasty of Korea as the first standardized rain gauge. In 1662, Christopher Wren created the first tipping-bucket rain gauge in Britain in collaboration with Robert Hooke. Hooke also designed a manual gauge with a funnel that made measurements throughout 1695.
Richard Towneley was the first to make systematic rainfall measurements over a period of 15 years from 1677 to 1694, publishing his records in the Philosophical Transactions of the Royal Society. Towneley called for more measurements elsewhere in the country to compare the rainfall in different regions, although only William Derham appears to have taken up Towneley's challenge. They jointly published the rainfall measurements for Towneley Park and Upminster in Essex for the years 1697 to 1704.
The naturalist Gilbert White took measurements to determine the mean rainfall from 1779 to 1786, although it was his brother-in-law, Thomas Barker, who made regular and meticulous measurements for 59 years, recording temperature, wind, barometric pressure, rainfall and clouds. His meteorological records are a valuable resource for knowledge of the 18th-century British climate. He was able to demonstrate that the average rainfall varied greatly from year to year with little discernible pattern.
The meteorologist George James Symons published the first annual volume of British Rainfall in 1860. This pioneering work contained rainfall records from 168 land stations in England and Wales. He was elected to the council of the British Meteorological Society in 1863 and made it his life's work to investigate rainfall within the British Isles. He set up a voluntary network of observers, who collected data which were returned to him for analysis. So successful was he in this endeavour that by 1866 he was able to show results that gave a fair representation of the distribution of rainfall and the number of recorders gradually increased until the last volume of British Rainfall which he lived to edit, for 1899, contained figures from 3,528 stations — 2,894 in England and Wales, 446 in Scotland, and 188 in Ireland. He also collected old rainfall records going back over a hundred years. In 1870 he produced an account of rainfall in the British Isles starting in 1725.
Due to the ever-increasing numbers of observers, standardisation of the gauges became necessary. Symons began experimenting with new gauges in his own garden. He tried different models with variations in size, shape, and height. In 1863 he began a collaboration with Michael Foster Ward from Calne, Wiltshire, who undertook more extensive investigations. By including Ward and various others around Britain, the investigations continued until 1890. The experiments were remarkable for their planning, execution, and drawing of conclusions. The results of these experiments led to the progressive adoption of the well-known standard gauge, still used by the UK Meteorological Office today, namely, one made of "... copper, with a five-inch funnel having its brass rim one foot above the ground ..."
Most modern rain gauges generally measure the precipitation in millimetres in height collected during a certain period, equivalent to litres per square metre. Previously rain was recorded as inches or points, where one point is equal to 0.254 mm or 0.01 of an inch.
Rain gauge amounts are read either manually or by automatic weather station (AWS). The frequency of readings will depend on the requirements of the collection agency. Some countries will supplement the paid weather observer with a network of volunteers to obtain precipitation data (and other types of weather) for sparsely populated areas.
In most cases the precipitation is not retained, but some stations do submit rainfall and snowfall for testing, which is done to obtain levels of pollutants.
Rain gauges have their limitations. Attempting to collect rain data in a tropical cyclone can be nearly impossible and unreliable (even if the equipment survives) due to wind extremes. Also, rain gauges only indicate rainfall in a localized area. For virtually any gauge, drops will stick to the sides or funnel of the collecting device, such that amounts are very slightly underestimated, and those of .01 inches or .25 mm may be recorded as a "trace".
Another problem encountered is when the temperature is close to or below freezing. Rain may fall on the funnel and ice or snow may collect in the gauge, blocking subsequent rain. To alleviate this, a gauge may be equipped with an automatic electric heater to keep its moisture-collecting surfaces and sensor slightly above freezing.
Rain gauges should be placed in an open area where there are no buildings, trees, or other obstacles to block the rain. This is also to prevent the water collected on the roofs of buildings or the leaves of trees from dripping into the rain gauge after a rain, resulting in inaccurate readings. Rain gauges can help people comprehend the amount of precipitation fallen down in a certain period of time.
Types
Types of rain gauges include graduated cylinders, weighing gauges, tipping bucket gauges, and simply buried pit collectors. Each type has its advantages and disadvantages while collecting rain data.
U.S. standard rain gauge
The standard United States National Weather Service rain gauge, developed at the start of the 20th century, consists of an 8 in (200 mm) funnel emptying into a metal graduated cylinder, 2.525 in (64.1 mm) in diameter, which fits inside a larger container that is 8 in (200 mm) in diameter and 20 in (510 mm) tall. If the rainwater overflows the graduated inner cylinder, then the larger outer container will catch it. When measurements are taken, then the height of the water in the small graduated cylinder is measured, and the excess overflow in the large container is carefully poured into another graduated cylinder and measured to give the total rainfall. A cone meter is sometimes used to prevent leakage that can result in alteration of the data. In locations using the metric system, the cylinder is usually marked in mm and will measure up to 250 mm (9.8 in) of rainfall.
Each horizontal line on the cylinder is 0.5 mm (0.02 in). In designs made for areas using Imperial units, each horizontal line represents 0.01 in (0.25 mm) inches.
Pluviometer of intensities
The pluviometer of intensities (or Jardi's pluviometer) is a tool that measures the average intensity of rainfall in a certain interval of time. It was initially designed to record the rainfall regime in Catalonia but eventually spread throughout the world.
It employs the principle of feedback ... the incoming water pushes the buoy upwards, making the lower "adjusting conic needle" to let pass the same amount of water that enters into the container, this way ... the needle records on the drum the amount of water flowing through it at every moment—in mm of rainfall per square meter.
It consists of a rotating drum that rotates at constant speed, this drum drags a graduated sheet of cardboard, which has the time at the abscissa while the y-axis indicates the height of rainfall in mm of rain. This height is recorded with a pen that moves vertically, driven by a buoy, marking on the paper the rainfall over time. Each cardboard sheet is usually used for one day.
As the rain falls, the water collected by the funnel falls into the container and raises the buoy that makes the pen arm rise in the vertical axis, marking the cardboard accordingly. If the rainfall does not vary, the water level in the container remains constant, and while the drum rotates, the pen's mark is more or less a horizontal line, proportional to the amount of water that has fallen. When the pen reaches the top edge of the recording paper, it means that the buoy is "up high in the tank" leaving the tip of the conical needle in a way that uncovers the regulating hole, i.e., the maximum flow that the apparatus is able to record. If the rain suddenly decreases, making the container (as it empties) quickly lower the buoy, that movement corresponds to a steep slope line that can reach the bottom of the recorded cardboard if it stops raining.
The rain gauge of intensities allowed precipitation to be recorded over many years, particularly in Barcelona (95 years), apart from many other places around the world, such as Hong Kong.
Weighing precipitation gauge
A weighing-type precipitation gauge consists of a storage bin, which is weighed to record the mass. Certain models measure the mass using a pen on a rotating drum, or by using a vibrating wire attached to a data logger. The advantages of this type of gauge over tipping buckets are that it does not underestimate intense rain, and it can measure other forms of precipitation, including rain, hail, and snow. These gauges are, however, more expensive and require more maintenance than tipping bucket gauges.
The weighing-type recording gauge may also contain a device to measure the number of chemicals contained in the location's atmosphere. This is extremely helpful for scientists studying the effects of greenhouse gases released into the atmosphere and their effects on the levels of the acid rain. Some Automated Surface Observing System (ASOS) units use an automated weighing gauge called the AWPAG (All Weather Precipitation Accumulation Gauge).
Tipping bucket rain gauge
The tipping bucket rain gauge consists of a funnel that collects and channels the precipitation into a small seesaw-like container. After a pre-set amount of precipitation falls, the lever tips, dumping the collected water and sending an electrical signal. An old-style recording device may consist of a pen mounted on an arm attached to a geared wheel that moves once with each signal sent from the collector. In this design, as the wheel turns the pen arm moves either up or down leaving a trace on the graph and at the same time making a loud "click".
The tipping bucket rain gauge is not as accurate as the standard rain gauge, because the rainfall may stop before the lever has tipped. When the next period of rain begins it may take no more than one or two drops to tip the lever. This would then indicate that a pre-set amount has fallen when only a fraction of that amount has actually fallen. Tipping buckets also tend to underestimate the amount of rainfall, particularly in snowfall and heavy rainfall events. The advantage of the tipping bucket rain gauge is that the character of the rain (light, medium, or heavy) may be easily obtained. Rainfall character is decided by the total amount of rain that has fallen in a set period (usually 1 hour) by counting the number of pulses during that period. Algorithms may be applied to the data as a method of correcting the data for high-intensity rainfall.
Modern tipping rain gauges consist of a plastic collector balanced over a pivot. When it tips, it actuates a switch (such as a reed switch) which is then electronically recorded or transmitted to a remote collection station.
Tipping gauges can also incorporate elements of weighing gauges whereby a strain gauge is fixed to the collection bucket so that the exact rainfall can be read at any moment. Each time the collector tips, the strain gauge (weight sensor) is re-zeroed to null out any drift.
To measure the water equivalent of frozen precipitation, a tipping bucket may be heated to melt any ice and snow that is caught in its funnel. Without a heating mechanism, the funnel often becomes clogged during a frozen precipitation event, and thus no precipitation can be measured. Many Automated Surface Observing System (ASOS) units use heated tipping buckets to measure precipitation.
Optical rain gauge
This type of gauge has a row of collection funnels. In an enclosed space below each is a laser diode and a photo transistor detector. When enough water is collected to make a single drop, it drops from the bottom, falling into the laser beam path. The sensor is set at right angles to the laser so that enough light is scattered to be detected as a sudden flash of lights. The flashes from these photodetectors are then read and transmitted or recorded. Different type of optical range gauges have been used throughout the decades. The technology has also improved.
Acoustic rain gauge
Acoustic disdrometers, also referred to as hydrophones, are able to sense the sound signatures for each drop size as rain strikes a water surface within the gauge. Since each sound signature is unique, it is possible to invert the underwater sound field to estimate the drop-size distribution within the rain. Selected moments of the drop-size distribution yield rainfall rate, rainfall accumulation, and other rainfall properties.
Additional Information
A rain gauge is a meteorological instrument to measure the precipitating rain in a given amount of time per unit area. The instrument consists of a collection container which is placed in an open area. The precipitation is measured in terms of the height of the precipitated water accumulated in the container per given time and is expressed in millimetres. Since the same amount of rain precipitation is assumed to be occurring around the container, the area of collection is not a factor. However, it should not be too small, neither should it be too large. Due to spatial uniformity of rainfall, 1 mm of measured precipitation is the equivalent of 1 L of precipitated rain water volume per metre squared.
A tipping bucket rain gauge consists of a pair of rainwater collecting buckets. It is covered by a funnel, with an open collector area at the top where A is the area of collection. The buckets are so placed on a pivot that only one bucket remains under the funnel at a time. During rain, rain water is collected in the collecting bucket, through the funnel. When the water fills up to a known point of the bucket, say having a volume v, the bucket tips, emptying the water. When one bucket tips, the other bucket quickly moves into place to collect rainwater. Each time a bucket tips, an electronic signal is sent to a recorder which is registered by the instrument with time stamp.
It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2597 2025-09-24 16:39:37
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2397) Mount Everest
Gist
Mount Everest is Earth's highest mountain above sea level, located in the Himalayas on the border of Nepal and China, with a peak elevation of 8,848.86 meters (29,031.7 ft). It is known by local names Sagarmatha (Nepali) and Chomolungma (Tibetan), and was first climbed by Sir Edmund Hillary and Tenzing Norgay in 1953. The mountain attracts experienced climbers and trekkers, with professional guides and Sherpas providing crucial local expertise for the challenging ascent.
Mount Everest is located on the border between two countries: Nepal and China. The summit straddles the international boundary, with its slopes in both the Solukhumbu district of Nepal and the Tibet Autonomous Region of China.
Summary
Mount Everest (known locally as Sagarmāthā in Nepal and Qomolangma in Tibet) is Earth's highest mountain above sea level. It lies in the Mahalangur Himal sub-range of the Himalayas and marks part of the China–Nepal border at its summit. Its height was most recently measured in 2020 by Chinese and Nepali authorities as 8,848.86 m (29,031 ft 8 1⁄2 in).
Mount Everest attracts many climbers, including highly experienced mountaineers. There are two main climbing routes, one approaching the summit from the southeast in Nepal (known as the standard route) and the other from the north in Tibet. While not posing substantial technical climbing challenges on the standard route, Everest presents dangers such as altitude sickness, weather, and wind, as well as hazards from avalanches and the Khumbu Icefall. As of May 2024, 340 people have died on Everest. Over 200 bodies remain on the mountain and have not been removed due to the dangerous conditions.
Climbers typically ascend only part of Mount Everest's elevation, as the mountain's full elevation is measured from the geoid, which approximates sea level. The closest sea to Mount Everest's summit is the Bay of Bengal, almost 700 km (430 mi) away. To approximate a climb of the entire height of Mount Everest, one would need to start from this coastline, a feat accomplished by Tim Macartney-Snape's team in 1990. Climbers usually begin their ascent from base camps above 5,000 m (16,404 ft). The amount of elevation climbed from below these camps varies. On the Tibetan side, most climbers drive directly to the North Base Camp. On the Nepalese side, climbers generally fly into Kathmandu, then Lukla, and trek to the South Base Camp, making the climb from Lukla to the summit about 6,000 m (20,000 ft) in elevation gain.
The first recorded efforts to reach Everest's summit were made by British mountaineers. As Nepal did not allow foreigners to enter the country at the time, the British made several attempts on the North Ridge route from the Tibetan side. After the first reconnaissance expedition by the British in 1921 reached 7,000 m (22,966 ft) on the North Col, the 1922 expedition on its first summit attempt marked the first time a human had climbed above 8,000 m (26,247 ft) and it also pushed the North Ridge route up to 8,321 m (27,300 ft). On the 1924 expedition George Mallory and Andrew Irvine made a final summit attempt on 8 June but never returned, sparking debate as to whether they were the first to reach the top. Tenzing Norgay and Edmund Hillary made the first documented ascent of Everest in 1953, using the Southeast Ridge route. Norgay had reached 8,595 m (28,199 ft) the previous year as a member of the 1952 Swiss expedition. The Chinese mountaineering team of Wang Fuzhou, Gonpo, and Qu Yinhua made the first reported ascent of the peak from the North Ridge on 25 May 1960.
Details
Mount Everest, mountain on the crest of the Great Himalayas of southern Asia that lies on the border between Nepal and the Tibet Autonomous Region of China, at 27°59′ N 86°56′ E. Reaching an elevation of 29,032 feet (8,849 meters), Mount Everest is the highest mountain in the world.
Like other high peaks in the region, Mount Everest has long been revered by local peoples. Its most common Tibetan name, Chomolungma, means “Goddess Mother of the World” or “Goddess of the Valley.” The Sanskrit name Sagarmatha means literally “Peak of Heaven.” Its identity as the highest point on the Earth’s surface was not recognized, however, until 1852, when the governmental Survey of India established that fact. In 1865 the mountain—previously referred to as Peak XV—was renamed for Sir George Everest, British surveyor general of India from 1830 to 1843. According to etymological sources, the mountain’s Western name is often mispronounced “Ever-est” or “Ev-rest,” despite the pronunciation of Sir George Everest’s surname being “EVE-rest.”
Physical features:
Geology and relief
The Himalayan ranges were thrust upward by tectonic action as the Indian-Australian Plate moved northward from the south and was subducted (forced downward) under the Eurasian Plate following the collision of the two plates between about 40 and 50 million years ago. The Himalayas themselves started rising about 25 to 30 million years ago, and the Great Himalayas began to take their present form during the Pleistocene Epoch (about 2,600,000 to 11,700 years ago). Everest and its surrounding peaks are part of a large mountain massif that forms a focal point, or knot, of this tectonic action in the Great Himalayas. Information from global positioning instruments in place on Everest since the late 1990s indicates that the mountain continues to move a few inches to the northeast and rise a fraction of an inch each year.
Everest is composed of multiple layers of rock folded back on themselves (nappes). Rock on the lower elevations of the mountain consists of metamorphic schists and gneisses, topped by igneous granites. Higher up are found sedimentary rocks of marine origin (remnants of the ancient floor of the Tethys Sea that closed after the collision of the two plates). Notable is the Yellow Band, a limestone formation that is prominently visible just below the summit pyramid.
The barren Southeast, Northeast, and West ridges culminate in the Everest summit; a short distance away is the South Summit, a minor bump on the Southeast Ridge with an elevation of 28,700 feet (8,748 meters). The mountain can be seen directly from its northeastern side, where it rises about 12,000 feet (3,600 meters) above the Plateau of Tibet. The peak of Changtse (24,803 feet [7,560 meters]) rises to the north. Khumbutse (21,867 feet [6,665 meters]), Nuptse (25,791 feet [7,861 meters]), and Lhotse (27,940 feet [8,516 meters]) surround Everest’s base to the west and south.
Everest is shaped like a three-sided pyramid. The three generally flat planes constituting the sides are called faces, and the line by which two faces join is known as a ridge. The North Face rises above Tibet and is bounded by the North Ridge (which meets the Northeast Ridge) and the West Ridge; key features of this side of the mountain include the Great and Hornbein couloirs (steep gullies) and the North Col at the start of the North Ridge. The Southwest Face rises above Nepal and is bounded by the West Ridge and the Southeast Ridge; notable features on this side include the South Col (at the start of the Southeast Ridge) and the Khumbu Icefall, the latter a jumble of large blocks of ice that has long been a daunting challenge for climbers. The East Face—or Kangshung (Kangxung) Face—also rises above Tibet and is bounded by the Southeast Ridge and the Northeast Ridge.
The summit of Everest itself is covered by rock-hard snow surmounted by a layer of softer snow that fluctuates annually by some 5–20 feet (1.5–6 meters); the snow level is highest in September, after the monsoon, and lowest in May after having been depleted by the strong northwesterly winter winds. The summit and upper slopes sit so high in the Earth’s atmosphere that the amount of breathable oxygen there is one-third what it is at sea level. Lack of oxygen, powerful winds, and extremely cold temperatures preclude the development of any plant or animal life there.
Drainage and climate
Glaciers cover the slopes of Everest to its base. Individual glaciers flanking the mountain are the Kangshung Glacier to the east; the East, Central, and West Rongbuk (Rongpu) glaciers to the north and northwest; the Pumori Glacier to the northwest; and the Khumbu Glacier to the west and south, which is fed by the glacier bed of the Western Cwm, an enclosed valley of ice between Everest and the Lhotse-Nuptse Ridge to the south. Glacial action has been the primary force behind the heavy and continuous erosion of Everest and the other high Himalayan peaks.
The mountain’s drainage pattern radiates to the southwest, north, and east. The Khumbu Glacier melts into the Lobujya (Lobuche) River of Nepal, which flows southward as the Imja River to its confluence with the Dudh Kosi River. In Tibet the Rong River originates from the Pumori and Rongbuk glaciers and the Kama River from the Kangshung Glacier: both flow into the Arun River, which cuts through the Himalayas into Nepal. The Rong, Dudh Kosi, and Kama river valleys form, respectively, the northern, southern, and eastern access routes to the summit.
The climate of Everest is always hostile to living things. The warmest average daytime temperature (in July) is only about −2 °F (−19 °C) on the summit; in January, the coldest month, summit temperatures average −33 °F (−36 °C) and can drop as low as −76 °F (−60 °C). Storms can come up suddenly, and temperatures can plummet unexpectedly. The peak of Everest is so high that it reaches the lower limit of the jet stream, and it can be buffeted by sustained winds of more than 100 miles (160 km) per hour. Precipitation falls as snow during the summer monsoon (late May to mid-September). The risk of frostbite to climbers on Everest is extremely high.
The height of Everest
Controversy over the exact elevation of the summit developed because of variations in snow level, gravity deviation, and light refraction. The figure 29,028 feet (8,848 meters), plus or minus a fraction, was established by the Survey of India between 1952 and 1954 and became widely accepted. This value was used by most researchers, mapping agencies, and publishers until 1999.
Attempts were subsequently made to remeasure the mountain’s height. A Chinese survey in 1975 obtained the figure of 29,029.24 feet (8,848.11 meters), and an Italian survey, using satellite surveying techniques, obtained a value of 29,108 feet (8,872 meters) in 1987, but questions arose about the methods used. In 1992 another Italian survey, using the Global Positioning System (GPS) and laser measurement technology, yielded the figure 29,023 feet (8,846 meters) by subtracting from the measured height 6.5 feet (2 meters) of ice and snow on the summit, but the methodology used was again called into question.
In 1999 an American survey, sponsored by the (U.S.) National Geographic Society and others, took precise measurements using GPS equipment. Their finding of 29,035 feet (8,850 meters), plus or minus 6.5 feet (2 meters), was accepted by the society and by various specialists in the fields of geodesy and cartography. The Chinese mounted another expedition in 2005 that utilized ice-penetrating radar in conjunction with GPS equipment. The result of this was what the Chinese called a “rock height” of 29,017.12 feet (8,844.43 meters), which, though widely reported in the media, was recognized only by China for the next several years. Nepal in particular disputed the Chinese figure, preferring what was termed the “snow height” of 29,028 feet. In April 2010 China and Nepal agreed to recognize the validity of both figures. In December 2020 China and Nepal jointly declared that the height of Everest was 29,031.69 feet (8,848.86 meters). This new measurement, derived from data from surveys performed by Nepal in 2019 and China in 2020 that utilized GPS and BeiDou navigation technology and laser theodolites, was accepted by various specialists in the fields of geodesy and cartography, including the National Geographic Society.
Human factors:
Habitation
Everest is so tall and its climate so severe that it is incapable of supporting sustained human occupation, but the valleys below the mountain are inhabited by Tibetan-speaking peoples. Notable among these are the Sherpas, who live in villages at elevations up to about 14,000 feet (4,270 meters) in the Khumbu valley of Nepal and other locations. Traditionally an agricultural people with little cultivable land at their disposal, the Sherpas for years were traders and led a seminomadic lifestyle in their search for pastureland. In summer, livestock was grazed as high as 16,000 feet (4,880 meters), while winter refuge was taken at lower elevations on sheltered ledges and along riverbanks.
Living in close proximity to the world’s highest mountains, the Sherpas traditionally treated the Himalayas as sacred—building Buddhist monasteries at their base, placing prayer flags on the slopes, and establishing sanctuaries for the wildlife of the valleys that included musk deer, monal pheasant, and Himalayan partridge. Gods and demons were believed to live in the high peaks, and the Yeti (the so-called Abominable Snowman) was said to roam the lower slopes. For these reasons, the Sherpas traditionally did not climb the mountains.
However, beginning with the British expeditions of the early 20th century, surveying and portering work became available. Eventually, the respect and pay earned in mountaineering made it attractive to the Sherpas, who, being so well adapted to the high altitudes, were capable of carrying large loads of cargo over long distances. Though Sherpas and other hill people (the name Sherpa came to be applied—erroneously—to all porters) tend to outperform their foreign clients, they typically have played a subordinate role in expeditions; rarely, for example, has one of their names been associated with a pioneering route on Everest. The influx of foreign climbers—and, in far greater numbers, trekkers—has dramatically changed Sherpa life, as their livelihood increasingly has come to depend on these climbing expeditions.
Additional Information
Mount Everest is a peak in the Himalaya mountain range. It is located between Nepal and Tibet, an autonomous region of China. At 8,849 meters (29,032 feet), it is considered the tallest point on Earth. In the nineteenth century, the mountain was named after George Everest, a former Surveyor General of India. The Tibetan name is Chomolungma, which means “Mother Goddess of the World.” The Nepali name is Sagarmatha, which has various meanings.
The first ever recorded people to climb Everest were Edmund Hillary (a mountaineer from New Zealand) and his Tibetan guide Tenzing Norgay. They climbed the mountain in 1953 and hold the record together. The first records of Everest’s height came much earlier, in 1856. British surveyors recorded that Everest was the tallest peak in the world in their Great Trigonometrical Survey of the Indian subcontinent.
The Himalayan mountains have long been home to indigenous groups living in the valleys. The most famous of these are the Sherpa people. The word “Sherpa” is often used to mean mountain guide, though it actually refers to an ethnic group. The Sherpa have valuable experience in mountain climbing, which they can provide to other climbers. Most climbs of Everest would be impossible without the Sherpas’ logistical help and knowledge. However, their way of life extends beyond helping Everest climbers. Traditionally, their lifestyle has consisted of farming, herding, and trade. And, because they live at such a high altitude year round, they are accustomed to the low oxygen levels.
Climbing Mount Everest has become a popular expedition for mountain climbers. However, it is a dangerous undertaking. Climbing Everest requires a lot of experience mountaineering elsewhere, as well as a certificate of good health, equipment, and a trained Nepalese guide. The snow and ice on the mountain create deadly hazards like avalanches, and there is only a limited climbing season due to bad weather conditions. But perhaps the biggest danger is the altitude. Most climbers are not accustomed to the high altitude and low oxygen levels and rely on bottled oxygen they bring along. This is why the area above 8,000 meters (26,000 feet) elevation on Everest is called the “death zone.” Climbers who spend long periods in this region can develop altitude sickness and even brain swelling.
Mount Everest’s climbing industry has become controversial. As popularity of the climb has increased, there have been more “traffic jams” as climbers spend too much time in the death zone waiting for their chance to go to the summit. With more people has also come more pollution up near the summit as climbers often discard unwanted items all along the mountain. Additionally, the Sherpa people have been exploited by climbers, and their traditional way of life has been disrupted by foreign climbers. Sherpa guides are faced with some of the highest death rates of any field of employment, for comparatively little pay. Most disturbingly, because many climbers have died along the way, and their bodies are impossible to retrieve, climbers must frequently travel past corpses as they make their way up the mountain.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2598 2025-09-25 18:03:13
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2398) Bee Hummingbird
Gist
The Bee Hummingbird is a specific type of hummingbird, notable for being the smallest bird in the world, while "hummingbird" refers to the entire family of small birds known for their hovering flight and nectar-feeding habits. The key differences are size and species, with the Bee Hummingbird being exceptionally tiny (around 2 inches long and weighing under 2 grams), whereas other hummingbirds vary in size but are generally larger.
Males grow to be 5.51 cm long and weigh 1.95 g, on average, while females grow to be 6.12 cm long and weigh 2.6 g, on average. This small species is very compact and agile with an average wingspan of 3.25 cm. These birds have straight and rather short beaks when compared with other species of hummingbirds.
The Smallest Living Bird
Yes, the bee hummingbird (Mellisuga helenae) is the smallest living bird in the world. Native to Cuba, this tiny bird is roughly the size of a bee, measuring about 5 centimeters (2 inches) long and weighing around 1.5 to 2 grams. Despite its small size, it is a powerful pollinator, visiting over 1,500 flowers a day for nectar.
Summary
Would you like to see the world’s smallest bird? Then you’ll need to travel to Cuba.
Once on the island, your best bet for tracking down the tiny wonder is to visit a forest edge hung heavily with vines and bromeliads. There, hovering at the flowers — if you squint hard enough — you’ll find the Bee Hummingbird.
The Bee Hummingbird, which is found only in Cuba, is an absolute miniature, even among hummingbirds. It measures a mere two and a quarter inches long. Bee Hummingbirds are often mistaken for bees. They weigh less than two grams — less than a dime. That’s half the weight of our backyard hummers, like the Ruby-throated or Rufous. The female builds a nest barely an inch across. Her eggs are about the size of a coffee bean.
In flight, the Bee Hummingbird’s tiny wings beat 80 times a second. And during a courtship flight, they beat up to 200 times per second! The male’s entire head and throat shine in fiery pinkish-red, and blazing red feathers point like spikes down the sides of the breast.
Details
The bee hummingbird, zunzuncito or Helena hummingbird (Mellisuga helenae) is a species of hummingbird, native to the island of Cuba in the Caribbean. It is the smallest known bird.[3][4] The bee hummingbird feeds on nectar of flowers and bugs found in Cuba.
Description
The bee hummingbird is the smallest living bird. Females weigh 2.6 g (0.092 oz) and are 6.1 cm (2+3/8 in) long, and are slightly larger than males, which have an average weight of 1.95 g (0.069 oz) and length of 5.5 cm (2+1⁄8 in). Like all hummingbirds, it is a swift, strong flier.
The male has a green pileum and bright red throat, iridescent gorget with elongated lateral plumes, bluish upper parts, and the rest of the underparts mostly greyish white. Compared to other small hummingbirds, which often have a slender appearance, the bee hummingbird looks rounded and plump.
Female bee hummingbirds are bluish green with a pale gray underside. The tips of their tail feathers have white spots. During the mating season, males have a reddish to pink head, chin, and throat. The female lays only two eggs at a time, each about the size of a coffee bean.
The bee hummingbird's feathers have iridescent colors, which is not always noticeable, but depends on the viewing angle. The bird's slender, pointed bill is adapted for probing deep into flowers. The bee hummingbird feeds mainly on nectar, by moving its tongue rapidly in and out of its mouth. In the process of feeding, the bird picks up pollen on its bill and head. When it flies from flower to flower, it transfers the pollen. In this way, it plays an important role in plant reproduction. In one day, the bee hummingbird may visit 1,500 flowers. It is a diurnal bird that can fly at 40–48 km/h (22–26 kn; 11–13 m/s), and it beats its wings 80–200 times per second, which allows it to remain stationary in the air to feed on flowers. The bee hummingbird lives up to seven years in the wild, and 10 years in captivity.
The bee hummingbird has also been described as the smallest dinosaur. This characterization is based upon the recognition that birds are, in fact, a living form of theropod dinosaurs (or, strictly speaking, avian dinosaurs), and no smaller bird or non-avian dinosaur has been found in the fossil record.
The call is described as a "high pitched, jumbled twitter". Within their territory a male will often sing atop the highest tree.
Diet
The bee hummingbird has been reported to visit ten plant species, nine of them native to Cuba.
They occasionally eat insects and spiders. In a typical day, bee hummingbirds will consume up to half their body weight in food.
Taxonomy
The closest evolutionary relative of the bee hummingbird is the vervain hummingbird (Mellisuga minima), the only other member of its genus. The habitats of the vervain hummingbird are in Cuba's neighboring islands, Hispaniola and Jamaica.
Habitat and distribution
The bee hummingbird is endemic to the entire Cuban archipelago, including the main island of Cuba and the Isla de la Juventud in the West Indies. In these regions bee hummingbirds generally live in areas of thick growth that contain lianas and epiphytes. Its population is fragmented; it is found in Cuba's mogote areas in Pinar del Río Province and more commonly in Zapata Swamp (Matanzas Province) and in eastern Cuba, with reference localities in Alexander Humboldt National Park and Baitiquirí Ecological Reserve (Guantánamo Province) and Gibara and Sierra Cristal (Holguín Province).
Additional Information
The Bee Hummingbird, found only in Cuba, is the smallest bird in the world. An absolute miniature, even among hummingbirds, it measures only two and a quarter inches long. Often mistaken for bees, they weigh less than a dime. The female builds a nest barely an inch across, and lays eggs about the size of a coffee bean.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2599 2025-09-26 17:13:03
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2399) Suez Canal
Gist
The Suez Canal is an artificial waterway in Egypt connecting the Mediterranean Sea to the Red Sea, forming the shortest maritime route between Europe and Asia and separating Africa from Asia. Completed in 1869 by a French company, the canal significantly reduces shipping travel time and costs by eliminating the need to sail around Africa.
The Suez Canal connects the Mediterranean Sea to the Red Sea, making it the shortest maritime route to Asia from Europe. Since its completion in 1869, it has become one of the world's most heavily used shipping lanes.
Summary
The Suez Canal is an artificial sea-level waterway in Egypt, connecting the Mediterranean Sea to the Red Sea through the Isthmus of Suez and dividing Africa and Asia (and by extension, the Sinai Peninsula from the rest of Egypt). It is the border between Africa and Asia. The 193.30-kilometre-long (120.11 mi) canal is a key trade route between Europe and Asia.
In 1858, French diplomat Ferdinand de Lesseps formed the Compagnie de Suez for the express purpose of building the canal. Construction of the canal lasted from 1859 to 1869. The canal officially opened on 17 November 1869. It offers vessels a direct route between the North Atlantic and northern Indian oceans via the Mediterranean Sea and the Red Sea, avoiding the South Atlantic and southern Indian oceans and reducing the journey distance from the Arabian Sea to London by approximately 8,900 kilometres (5,500 mi), to 10 days at 20 knots (37 km/h; 23 mph) or 8 days at 24 knots (44 km/h; 28 mph). The canal extends from the northern terminus of Port Said to the southern terminus of Port Tewfik at the city of Suez. In 2021, more than 20,600 vessels traversed the canal (an average of 56 per day).
The original canal featured a single-lane waterway with passing locations in the Ballah Bypass and the Great Bitter Lake. It contained, according to Alois Negrelli's plans, no locks, with seawater flowing freely through it. In general, the water in the canal north of the Bitter Lakes flows north in winter and south in summer. South of the lakes, the current changes with the tide at Suez.
The canal was the property of the Egyptian government, but European shareholders, mostly British and French, owned the concessionary company which operated it until July 1956, when President Gamal Abdel Nasser nationalised it—an event which led to the Suez Crisis of October–November 1956. The canal is operated and maintained by the state-owned Suez Canal Authority (SCA) of Egypt. Under the Convention of Constantinople, it may be used "in time of war as in time of peace, by every vessel of commerce or of war, without distinction of flag." Nevertheless, the canal has played an important military strategic role as a naval short-cut and choke point. Navies with coastlines and bases on both the Mediterranean Sea and the Red Sea (Egypt and Israel) have a particular interest in the Suez Canal. After Egypt closed the Suez Canal at the beginning of the Six-Day War on 5 June 1967, the canal remained closed for eight years, reopening on 5 June 1975.
The Egyptian government launched construction in 2014 to expand and widen the Ballah Bypass for 35 km (22 mi) to speed up the canal's transit time. The expansion intended to nearly double the capacity of the Suez Canal, from 49 to 97 ships per day.[9] At a cost of LE 59.4 billion (US$9 billion), this project was funded with interest-bearing investment certificates issued exclusively to Egyptian entities and individuals.
The Suez Canal Authority officially opened the new side channel in 2016. This side channel, at the northern side of the east extension of the Suez Canal, serves the East Terminal for berthing and unberthing vessels from the terminal. As the East Container Terminal is located on the Canal itself, before the construction of the new side channel it was not possible to berth or unberth vessels at the terminal while a convoy was running.
Details
Suez Canal is a sea-level waterway running north-south across the Isthmus of Suez in Egypt to connect the Mediterranean and the Red seas. The canal separates the African continent from Asia, and it provides the shortest maritime route between Europe and the lands lying around the Indian and western Pacific oceans. It is one of the world’s most heavily used shipping lanes. The canal extends 193 km (120 miles) between Port Said (Būr Saʿīd) in the north and Suez in the south, with dredged approach channels north of Port Said, into the Mediterranean, and south of Suez. The canal does not take the shortest route across the isthmus, which is only 121 km (75 miles). Instead, it utilizes several lakes: from north to south, Lake Manzala (Buḥayrat al-Manzilah), Lake Timsah (Buḥayrat al-Timsāḥ), and the Bitter Lakes—Great Bitter Lake (Al-Buḥayrah al-Murrah al-Kubrā) and Little Bitter Lake (Al-Buḥayrah al-Murrah al-Ṣughrā). The Suez Canal is an open cut, without locks, and, though extensive straight lengths occur, there are eight major bends. To the west of the canal is the low-lying delta of the Nile River, and to the east is the higher, rugged, and arid Sinai Peninsula. Prior to construction of the canal (completed in 1869), the only important settlement was Suez, which in 1859 had 3,000 to 4,000 inhabitants. The rest of the towns along its banks have grown up since, with the possible exception of Al-Qanṭarah.
Physical features:
Geology
The Isthmus of Suez, the sole land bridge between the continents of Africa and Asia, is of relatively recent geologic origin. Both continents once formed a single large continental mass, but during the Paleogene and Neogene periods (about 66 to 2.6 million years ago) the great fault structures of the Red Sea and Gulf of Aqaba developed, with the opening and subsequent drowning of the Red Sea trough as far as the Gulf of Suez and the Gulf of Aqaba. In the succeeding Quaternary Period (about the past 2.6 million years), there was considerable oscillation of sea level, leading finally to the emergence of a low-lying isthmus that broadened northward to a low-lying open coastal plain. There the Nile delta once extended farther east—as a result of periods of abundant rainfall coincident with the Pleistocene Epoch (2,588,000 to 11,700 years ago)—and two river arms, or distributaries, formerly crossed the northern isthmus, one branch reaching the Mediterranean Sea at the narrowest point of the isthmus and the other entering the sea some 14.5 km (9 miles) east of present Port Said.
Physiography
Topographically, the Isthmus of Suez is not uniform. There are three shallow water-filled depressions: Lake Manzala, Lake Timsah, and the Bitter Lakes; though distinguished as Great and Little, the Bitter Lakes form one continuous sheet of water. A number of more-resistant bands of limestone and gypsum obtrude in the south of the isthmus, and another significant feature is a narrow valley leading from Lake Timsah southwestward toward the middle Nile delta and Cairo. The isthmus is composed of marine sediments, coarser sands, and gravels deposited in the early periods of abundant rainfall, Nile alluvium (especially to the north), and windblown sands.
When first opened in 1869, the canal consisted of a channel barely 8 metres (26 feet) deep, 22 metres (72 feet) wide at the bottom, and 61 to 91 metres (200 to 300 feet) wide at the surface. To allow ships to pass each other, passing bays were built every 8 to 10 km (5 to 6 miles). Construction involved the excavation and dredging of 74 million cubic metres (97 million cubic yards) of sediments. Between 1870 and 1884 some 3,000 groundings of ships occurred because of the narrowness and tortuousness of the channel. Major improvements began in 1876, and, after successive widenings and deepenings, the canal by the 1960s had a minimum width of 55 metres (179 feet) at a depth of 10 metres (33 feet) along its banks and a channel depth of 12 metres (40 feet) at low tide. Also in that period, passing bays were greatly enlarged and new bays constructed, bypasses were made in the Bitter Lakes and at Al-Ballāḥ, stone or cement cladding and steel piling for bank protection were almost entirely completed in areas particularly liable to erosion, tanker anchorages were deepened in Lake Timsah, and new berths were dug at Port Said to facilitate the grouping of ships in convoy.
Plans that had been made in 1964 for further enlargement were overtaken by the Arab-Israeli war of June 1967, during which the canal was blocked. The canal remained inoperative until June 1975, when it was reopened and improvements were recommenced. In 2015 the Egyptian government finished a nearly $8.5 billion project to upgrade the canal and significantly increase its capacity; nearly 29 km (18 miles) were added to its original length of 164 km (102 miles).
Additional Information
The Suez Canal is a man-made waterway that connects the Mediterranean Sea and the Red Sea. It runs north-south across a narrow piece of land called the Isthmus of Suez in Egypt. The canal separates Africa from Asia. It provides the shortest water route between Europe and the lands around the Indian and western Pacific oceans. The Suez Canal is one of the world’s most heavily used shipping lanes.
The canal extends 120 miles (193 kilometers) between the Egyptian cities of Port Said in the north and Suez in the south. The Isthmus of Suez is actually only 75 miles (121 kilometers) wide. However, the canal takes a longer route because it passes through several lakes. These are, from north to south, Lake Manzala, Lake Timsah, and the Bitter Lakes (Great Bitter Lake and Little Bitter Lake).
The Suez Canal crosses flat, sea-level terrain and requires no locks. Although extensive straight lengths occur, there are eight major bends. To the west of the canal is the low-lying delta of the Nile River. To the east is the higher, rugged, and arid Sinai Peninsula. The canal was begun in 1859 and completed 10 years later. Since then several towns have grown along its banks.
Physical Features
When first opened in 1869, the canal consisted of a channel barely 26 feet (8 meters) deep. It was 72 feet (22 meters) wide at the bottom and 200 to 300 feet (61 to 91 meters) wide at the surface. To allow ships to pass each other, passing bays were built every 5 to 6 miles (8 to 10 kilometers).
Construction involved the excavation and dredging of a huge amount of sediment. Between 1870 and 1884 some 3,000 ships became grounded because the canal was narrow and winding. Major improvements to the canal’s width and depth began in 1876.
By the 1960s the canal had a minimum width of 179 feet (55 meters) at a depth of 33 feet (10 meters). The channel depth was 40 feet (12 meters) at low tide. Also in that period, passing bays were enlarged and new bays were constructed. Workers reinforced most of the banks with stone, cement, or steel to help prevent erosion.
The next significant improvements were made in the 21st century. In 2015 the Egyptian government finished a nearly $8.5 billion project to upgrade the canal and significantly increase its capacity. The project added nearly 18 miles (29 kilometers) to its original length of 102 miles (164 kilometers).
Operation
The canal had its first full year of operation in 1870. That year ships used the canal 486 times, which is fewer than 2 ships per day. In 1966 there were 21,250 transits, for an average of 58 per day. The ships’ net tonnage, or the space available on all the ships, was about 274,000,000 long tons (278,400,000 metric tons). In 2014 there were fewer transits—only 17,148—but the net tonnage was about 947,800,000 long tons (963,000,000 metric tons).
The original canal did not permit two-way traffic. Ships needed to stop in a passing bay to allow the passage of ships in the other direction. It took a ship an average of 40 hours to pass through the canal. By 1939 the time had been drastically reduced. A system of ship convoys was adopted in 1947, consisting of one northbound and two southbound convoys per day. With that change and some enlargement work on the canal, transit time in the late 20th century ranged from 11 to 16 hours. After the 2015 improvements, northbound and southbound trips were both at about 11 hours each. The convoys travel the canal during both day and night.
To use the canal, ship owners must pay tolls, which are based on the weight and type of the cargo. Passengers have ridden without charge since 1950. As ships enter the canal at Port Said or Suez, workers assess them for tonnage and cargo. Each ship may pay as much as hundreds of thousands of dollars to travel through the canal. One or two Egyptian pilots board the ship to handle the actual canal transit. They may be replaced with new pilots at certain sections along the route. Radar plays an increasingly active role in their jobs.
In the early 21st century, cargo ships accounted for nearly all the canal traffic. The major northbound cargoes consist of crude petroleum and petroleum products, coal, ores and metals, and fabricated metals, as well as wood, oilseeds, and cereals. Southbound traffic consists of cement, fertilizers, fabricated metals, cereals, and empty oil tankers.
History
The first canal in the region is thought to have been dug as an irrigation channel about 1850 bc. It was known as the Canal of the Pharaohs. Subsequent rulers, including the Ptolemies and the Romans, expanded the canal. Historians believe that these early inhabitants used the canal to make trade easier from the delta lands to the Red Sea. In ad 775 the ʿAbbasid caliphs deliberately filled in the canal for military reasons.
European Involvement
The French made the first survey of the isthmus when they occupied Egypt in 1798–1801. French general Napoleon personally investigated the remains of the ancient canal and wanted to build a new one. However, a French engineer mistakenly calculated that the level of the Red Sea was higher than the Mediterranean. He reported that locks would be needed. The French subsequently suspended all work on a canal.
In 1854 the governor of Egypt, Saʿid Pasha, bestowed upon French diplomat Ferdinand de Lesseps an Act of Concession to construct a canal. In 1856 a second act gave the Suez Canal Company—jointly owned by the French and the Egyptians—the right to operate a maritime canal for 99 years after completion of the work. After that time, the Egyptian government would gain control of the canal. Construction began in 1859.
At first, authorities forced peasants to dig the canal with picks. Later, European laborers operating dredgers and steam shovels took over. Dredging was cheaper than dry excavation, so workers artificially flooded the terrain and dredged wherever possible. In August 1869 the waterway was completed. It was officially opened on November 17. Construction of the canal led to the growth of settlements in what had been almost uninhabited desert territory.
In 1875 financial troubles forced the new governor, Ismaʿil Pasha, to sell Egypt’s shares in the Suez Canal Company. The British government bought them, and France and Great Britain jointly controlled the canal for many years. In 1949 the company reinstated Egypt as a board member and gave the country some of the profits. It was also agreed that Egyptians would fill most new clerical jobs and technical appointments. In addition, the company would provide hospitals, schools, and other amenities for the workers.
Suez Crises
By the Convention of Constantinople (1888), the canal was to be an international waterway open to all countries in times of war and peace. This convention has not always been observed. During both world wars Germany and its allies attacked but failed to capture the canal. In 1949 Egypt denied the use of the canal to the Israelis and to all ships trading with Israel. That act followed Egypt’s and Israel’s involvement in the 1948 Arab-Israeli War.
In July 1956 Egypt’s President Gamal Abdel Nasser declared the canal to be Egyptian property. Since then an independent Egyptian government agency called the Suez Canal Authority has managed and operated the canal. In 1963 Egypt paid the last compensation to Suez Canal Company shareholders and became sole owner of the waterway.
The canal was closed to shipping twice in the 20th century. The first time was during the Suez Crisis of 1956–57. Nasser set off the crisis when he took over the canal in 1956, before the 99 years of the Suez Canal Company’s control were up. He did so after Britain and the United States refused to give Egypt funds they had previously promised for the construction of the Aswan High Dam. Nasser seized control of the canal so he could use its proceeds to pay for the dam. Diplomatic efforts to settle the crisis failed. Britain and France feared that Nasser might close the canal and cut off shipments of petroleum flowing from the Persian Gulf to western Europe. Their ally Israel invaded Egypt. French and British troops then occupied part of the canal zone. Several ships were trapped within the canal during that blockade. They were unable to leave until tensions eased and the canal was reopened in January 1957.
The second closing of the canal began in June 1967 during an Arab-Israeli war known as the Six-Day War. Egypt physically blocked both ends of the canal, trapping 15 ships in the canal’s Great Bitter Lake. In June 1975 Egypt reopened the canal. Four years later Egypt and Israel signed a peace treaty. Egypt again gave all ships access to the waterway.
21st Century
In 2014 Egyptian President Abdel Fattah al-Sisi launched a canal expansion project to boost Egypt’s economy. In August 2015 the government opened a new 22-mile (35-kilometer) expansion canal running parallel to the main channel. These separate channels allowed northbound and southbound ships to travel through that section without any delays. The main channel was deepened to allow for the passage of larger ships.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline
#2600 2025-09-27 17:22:05
- Jai Ganesh
- Administrator
- Registered: 2005-06-28
- Posts: 52,569
Re: Miscellany
2400) Panama Canal
Gist
The Panama Canal is an 82-kilometer (51-mile) artificial waterway in Panama that connects the Atlantic and Pacific oceans, saving ships from the long journey around South America. Owned and operated by Panama, it features a lock system to lift and lower vessels, and the 2016 expansion allows larger ships to pass through. This vital maritime route serves as a major conduit for global trade and revenue for Panama.
The Panama Canal serves as a maritime shortcut that saves time and costs in transporting all kinds of goods. The 80-kilometer waterway communicates the Atlantic and Pacific Oceans in one of the narrowest points of the American Continent.
Summary
The Panama Canal (Spanish: Canal de Panamá) is an artificial 82-kilometer (51-mile) waterway in Panama that connects the Caribbean Sea with the Pacific Ocean. It cuts across the narrowest point of the Isthmus of Panama, and is a conduit for maritime trade between the Atlantic and Pacific Oceans. Locks at each end lift ships up to Gatun Lake, an artificial fresh water lake 26 meters (85 ft) above sea level, created by damming the Chagres River and Lake Alajuela to reduce the amount of excavation work required for the canal. Locks then lower the ships at the other end. An average of 200,000,000 litres (52 million US gallons) of fresh water is used in a single passing of a ship. The canal is threatened by low water levels during droughts.
The Panama Canal shortcut greatly reduces the time for ships to travel between the Atlantic and Pacific oceans, enabling them to avoid the lengthy, hazardous route around the southernmost tip of South America via the Drake Passage, the Strait of Magellan or the Beagle Channel. Its construction was one of the largest and most difficult engineering projects ever undertaken. Since its inauguration on 15 August 1914, the canal has succeeded in shortening maritime communication in time and distance, invigorating maritime and economic transportation by providing a short and relatively inexpensive transit route between the two oceans, decisively influencing global trade patterns, boosting economic growth in developed and developing countries, as well as providing the basic impetus for economic expansion in many remote regions of the world.
Colombia, France, and later the United States controlled the territory surrounding the canal during construction. France began work on the canal in 1881, but stopped in 1889 because of a lack of investors' confidence due to engineering problems and a high worker mortality rate. The US took over the project in 1904 and opened the canal in 1914. The US continued to control the canal and surrounding Panama Canal Zone until the Torrijos–Carter Treaties provided for its handover to Panama in 1977. After a period of joint American–Panamanian control, the Panamanian government took control in 1999. It is now managed and operated by the Panamanian government-owned Panama Canal Authority.
The original locks are 33.5 meters (110 ft) wide and allow the passage of Panamax ships. A third, wider lane of locks was constructed between September 2007 and May 2016. The expanded waterway began commercial operation on 26 June 2016. The new locks allow for the transit of larger, Neopanamax ships.
Annual traffic has risen from about 1,000 ships in 1914, when the canal opened, to 14,702 vessels in 2008, for a total of 333.7 million Panama Canal/Universal Measurement System (PC/UMS) tons. By 2012, more than 815,000 vessels had passed through the canal. In that year, the top five users of the canal were the United States, China, Chile, Japan, and South Korea. In 2017, it took ships an average of 11.38 hours to pass between the canal's two outer locks. The American Society of Civil Engineers has ranked the Panama Canal one of the Seven Wonders of the Modern World.
Details
Panama Canal, lock-type canal, owned and administered by the Republic of Panama, that connects the Atlantic and Pacific oceans through the narrow Isthmus of Panama. The length of the Panama Canal from shoreline to shoreline is about 40 miles (65 km) and from deep water in the Atlantic (more specifically, the Caribbean Sea) to deep water in the Pacific about 50 miles (82 km). The canal, which was completed in August 1914, is one of the two most strategic artificial waterways in the world, the other being the Suez Canal. Ships sailing between the east and west coasts of the United States, which otherwise would be obliged to round Cape Horn in South America, shorten their voyage by about 8,000 nautical miles (15,000 km) by using the canal. Savings of up to 3,500 nautical miles (6,500 km) are also made on voyages between one coast of North America and ports on the other side of South America. Ships sailing between Europe and East Asia or Australia can save as much as 2,000 nautical miles (3,700 km) by using the canal.
From its opening in 1914 until 1979, the Panama Canal was controlled solely by the United States, which built it. In 1979, however, control of the canal passed to the Panama Canal Commission, a joint agency of the United States and the Republic of Panama, and complete control passed to Panama at noon on December 31, 1999. Administration of the canal is the responsibility of the Panama Canal Authority (Spanish: Autoridad del Canal de Panamá [ACP]), which answers solely to the government of Panama.
Physical features:
The canal
The Panama Canal lies at a latitude of 9° N, at a point where the North American Continental Divide dips to one of its lowest points. The canal does not, as is generally supposed, cross the isthmus from east to west. It runs due south from its entrance at Colón on the Atlantic side through the Gatún Locks to a point in the widest portion of Gatún Lake; it then turns sharply toward the east and follows a course generally to the southeast until it reaches the Bay of Panama, on the Pacific side. Its terminus near Balboa is some 25 miles (40 km) east of its terminus near Colón. Parallel to the canal are the Panama Canal Railway and the Boyd-Roosevelt Highway.
In passing from the Atlantic to the Pacific, vessels enter the approach channel in Limón Bay, which extends a distance of about 7 miles (11 km) to the Gatún Locks. At Gatún a series of three locks lifts vessels 85 feet (26 meters) to Gatún Lake. The lake, formed by Gatún Dam on the Chagres River and supplemented by waters from Alajuela Lake (Lake Madden; formed by the Madden Dam), covers an area of 166 square miles (430 square km). The channel through the lake varies in depth from 46 to 85 feet (14 to 26 meters) and extends for about 23 miles (37 km) to Gamboa. Gaillard (Culebra) Cut begins at Gamboa and passes through the Continental Divide. The channel through the cut has an average depth of about 43 feet (13 meters) and extends some 8 miles (13 km) to the Pedro Miguel Locks. The locks lower vessels 30 feet (9 meters) to Miraflores Lake, at an elevation of 52 feet (16 meters) above sea level. Vessels then pass through a channel almost 1.2 miles (2 km) long to the two-stepped locks at Miraflores, where they are lowered to sea level. The final segment of the canal is a dredged approach passage 7 miles long through which ships pass into the Pacific. Throughout its length the canal has a minimum bottom width of 500 feet (150 meters); in Gatún Lake the width of the channel varies between 500 and 1,000 feet (150 and 300 meters), and in Miraflores Lake the width is 740 feet (225 meters).
Locks
The canal locks operate by gravity flow of water from Gatún, Alajuela, and Miraflores lakes, which are fed by the Chagres and other rivers. The locks themselves are of uniform length, width, and depth and were built in pairs to permit the simultaneous transit of vessels in either direction. Each lock gate has two leaves, 65 feet (20 meters) wide and 6.5 feet (2 meters) thick, set on hinges. The gates range in height from 46 to 82 feet (14 to 25 meters); their movement is powered by electric motors recessed in the lock walls. They are operated from a control tower, which is located on the wall that separates each pair of locks and from which the flooding or emptying of the lock chambers is also controlled. The lock chambers are 1,000 feet (300 meters) long, 110 feet (33 meters) wide, and 40 feet (12 meters) deep.
Because of the delicate nature of the original lock mechanisms, only small craft are allowed to pass through the locks unassisted. Larger craft are guided by electric towing locomotives, which operate on cog tracks on the lock walls and serve to keep the ships centered in the lock. Before a lock can be entered, a fender chain, stretched between the walls of the approach, must be passed. If all is proceeding properly, that chain will be dropped into its groove at the bottom of the channel. If by any chance the ship is moving too rapidly for safety, the chain will remain stretched and the vessel will run against it. The chain, which is operated by hydraulic machinery in the walls, then will pay out slowly by automatic release until the vessel has been brought to a stop. If the vessel should get away from the towing locomotive and, breaking through the chain, ram the first gate, a second gate 50 feet (15 meters) away will protect the lock and arrest further advance.
The third lock systems of the Third Set of Locks Project, begun in 2007, were inspired by the Berendrecht lock in Antwerp, Belgium, and water-saving basins used in canals in Germany. Some 190,000 tons of steel, mostly from Mexico, were entrenched in heavily reinforced concrete to build the lock chambers on the Atlantic and Pacific sides, and the new lock gates measure up to 33 feet (10 meters) wide, 98 feet (30 meters) high, and 190 feet (58 meters) long. The new chambers and basins, which control the water flowing from Gatún Lake, were designed to minimize the turbulence of water flow and the disturbance to transiting vessels. The basins were completed in June 2016 and include 158 valves consisting of 20,000 tons of structural material. Officials say those water-saving basins are the largest in the world and facilitate a 60 percent reuse of water. Whereas the existing locks use 52 million gallons (197 million liters) with each use, the new locks use 48 million gallons (182 million liters).
Breakwaters
Long breakwaters have been constructed near the approach channels in both oceans. Breakwaters extend from the west and east sides of Limón Bay; the west breakwater protects the harbor against severe gales, and the east one reduces silting in the canal channel. On the Pacific side a causeway extends from Balboa to three small islands (Naos, Perico, and Flamenco) and diverts crosscurrents that carry soft material from the shallow harbor of Panama City into the canal channel.
Additional Information
The Panama Canal is approximately 80 kilometers long and connects the Atlantic and Pacific Oceans. This waterway was cut through one of the narrowest saddles of the isthmus that joins North and South America. The Canal uses a system of locks compartments with entrance and exit gates. The locks function as water lifts: they raise ships from sea level (the Pacific or the Atlantic) to the level of Gatun Lake (26 meters above sea level). Then, ships sail the channel through the Continental Divide.
Each set of locks bears the name of the town where it was built: Gatun (on the Atlantic side), and Pedro Miguel and Miraflores (on the Pacific side). The lock chambers -stepsare 33.53 meters wide by 304.8 meters long. The maximum dimensions of ships that can transit the Canal are: 32.3 meters in beam; draft -their depth reach- 12 meters in Tropical Fresh Water; and 294.1 meters long (depending on the type of ship).
The water used to raise and lower vessels in each set of locks flows from Gatun Lake by gravity; it comes into the locks through a system of main culverts that extend under the lock chambers from the sidewalls and the center wall.
The narrowest portion of the Canal is Culebra Cut, which extends from the north end of Pedro Miguel Locks to the south edge of Gatun Lake at Gamboa. This segment, approximately 13.7 kilometers long, is carved through the rock and shale of the Continental Divide.
Ships from all parts of the world transit daily through the Panama Canal. About 13 to 14 thousand vessels use the Canal every year. In fact, commercial transportation activities through the Canal represent approximately 5% of the world trade. The Canal has a work force of approximately 9 thousand employees and operates 24 hours a day, 365 days a year.
When the construction of new locks is completed, the canal will handle post-Panamax ships of maximum dimensions being: Length 365.8m, Breadth 48.8m, Draught 15.2m, with TEU capacity of 13,000.

It appears to me that if one wants to make progress in mathematics, one should study the masters and not the pupils. - Niels Henrik Abel.
Nothing is better than reading and gaining more and more knowledge - Stephen William Hawking.
Offline